Article Text

Statistics from Altmetric.com
- Age
- CD8 T cell
- Epstein–Barr virus
- multiple sclerosis
- T cell exhaustion, immunology
- virology
- neuromuscular
- EMG (single fibre)
- HMSN (Charcot-Marie-tooth)
- immunology
- Lambert Eaton syndrome
- channels
- neurophysiology
- neuromuscular
- motor neuron disease
- neuropathy
- neuromuscular
- virology
A large body of evidence indicates that infection with the Epstein–Barr virus (EBV) has a role in the pathogenesis of multiple sclerosis (MS).1 We have previously hypothesised that a genetically determined defect in the elimination of EBV-infected B cells by cytotoxic CD8 T cells might predispose to the development of MS by allowing EBV-infected autoreactive B cells to accumulate in the central nervous system (CNS).1 2 Recently, we have shown that patients with MS have a decreased frequency of CD8 T cells reactive to their own EBV-infected B cells.3 Since 1980, it has been recognised that MS patients have a decreased proportion and number of CD8 T cells in peripheral blood.4 This was initially interpreted as a decrease in CD8 suppressor T cells leading to disinhibition of autoimmune responses but was later attributed to sequestration of CD8 T cells in the CNS. An alternative explanation is that the CD8 T cell deficiency is genetically determined and causes the decreased CD8 T cell response to EBV,3 which allows the accumulation of EBV-infected B cells in the CNS and the consequent development of MS. In the present study, we have used flow cytometry to determine the frequency of CD8 T cells in the blood and its relationship to the EBV-specific T cell response and clinical features of MS.
Patients and methods
Blood was collected from 64 MS patients and 68 age- and sex-matched healthy subjects after obtaining informed consent. This study was approved by the Royal Brisbane & Women's Hospital Human Research Ethics Committee and The University of Queensland Medical Research Ethics Committee. All patients met the 2005 Revised McDonald Criteria for a diagnosis of MS. The clinical course was relapsing–remitting (RRMS) in 23 patients, secondary progressive MS (SPMS) in 25 patients and primary progressive MS (PPMS) in 16 patients. The patients had not received corticosteroids or immunomodulatory therapy for at least 3 months prior to venesection. Only two patients had ever received immunosuppressive drugs and these had been ceased 4 years before blood collection. An additional seven patients had received interferon-β, which had been ceased 6 months to 6 years before blood collection.
Peripheral blood mononuclear cells (PBMCs) were separated by density centrifugation and cryopreserved, as previously described.3 For flow cytometry, PBMC samples were thawed and cultured for 24 h to allow cells to rest and re-express cell surface receptors. PBMC samples were assessed using a Becton Dickinson FACSCalibur flow cytometer to determine the percentages of CD3 T cells, CD4CD3 T cells and CD8CD3 T cells within the combined lymphocyte and monocyte gates.
To measure the total T cell response to EBV-infected B cells, we performed interferon-γ (IFN-γ) enzyme-linked immunospot assays to determine the frequency of PBMC producing IFN-γ in response to autologous EBV-infected B cell lymphoblastoid cell lines (LCLs), as previously described.3 These assays were performed on blood samples that had been collected at the same time as the samples used for flow cytometry. CD8 T cells are the predominant population responding to LCLs in this enzyme-linked immunospot assay, with the frequency of EBV-specific T cells being five- to sevenfold higher in the CD8 population than in the CD4 population.3
Results
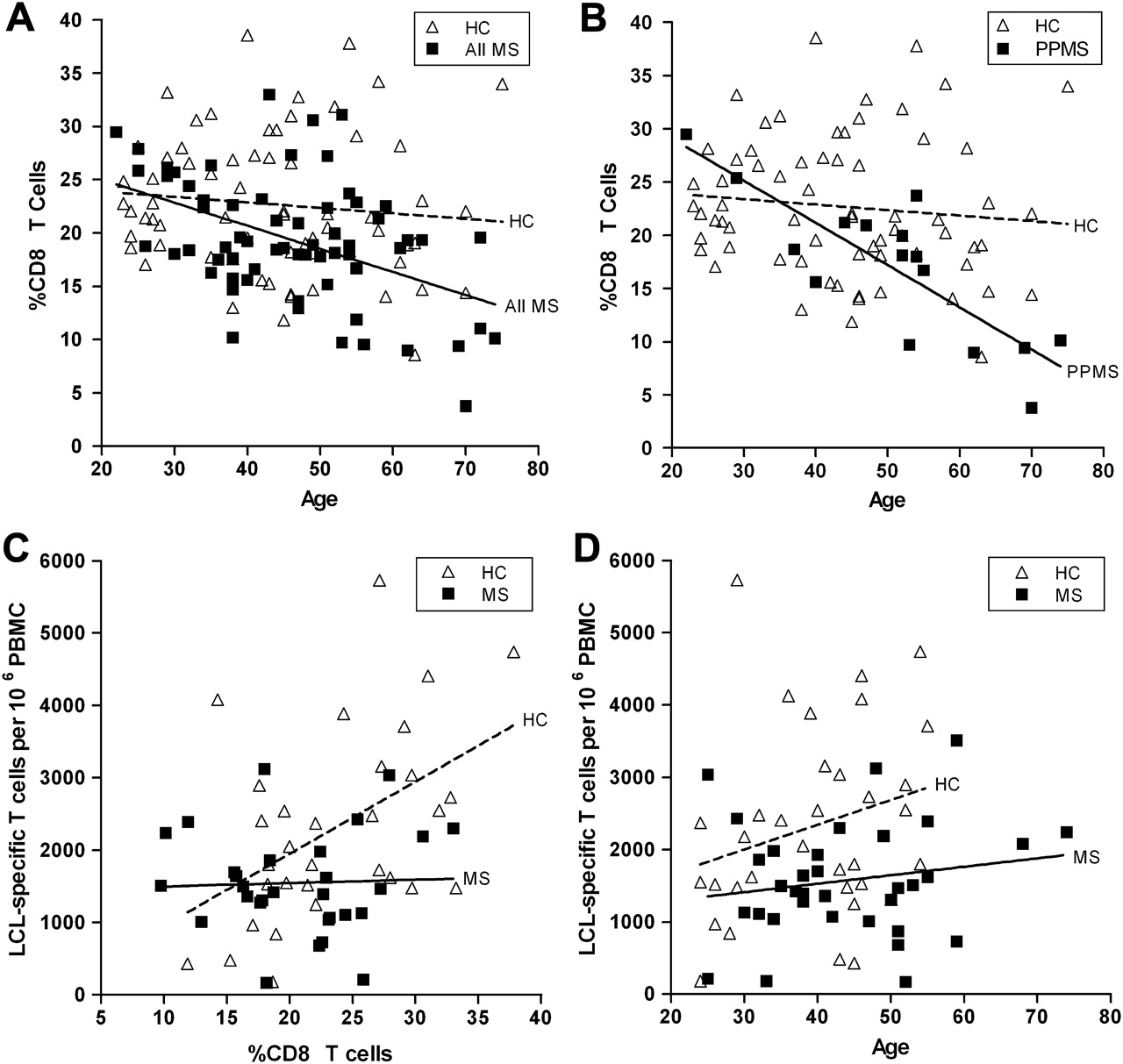
The mean percentage±SE of CD8 T cells was significantly decreased in MS patients (19.4±0.7) compared with healthy subjects (22.7±0.8, p<0.01, Mann–Whitney). The decrease in CD8 T cells was more pronounced in SPMS (18.8±1.0) and PPMS (16.9±1.7) than in RRMS (21.7±1.1). Strikingly, the proportion of CD8 T cells declined markedly with age in MS patients compared with healthy subjects (figure 1A). This decline was most marked in PPMS (figure1B). Multiple linear regression analysis demonstrated that patient age rather than Kurtzke Expanded Disability Status scale score or disease duration was the main determinant of the proportion of CD8 T cells in MS patients (p<0.05).

{kind=link}
(A) Proportion of CD8 T cells in peripheral blood mononuclear cells (PBMCs) in healthy control (HC) subjects and the total group of multiple sclerosis patients (All MS) plotted against age in years. The proportion of CD8 T cells declines markedly with increasing age in the patients with MS (continuous line) (r=−0.46, p=0.0001, Pearson) but not in the healthy subjects (broken line) (rs=−0.11, p=0.26, Spearman). The slopes of the regression lines are significantly different (p<0.05, multiple linear regression). (B) Proportion of CD8 T cells in PBMCs in HC subjects and patients with primary progressive MS (PPMS) plotted against age in years. The proportion of CD8 T cells declines markedly with age in patients with PPMS (continuous line) (r=−0.82, p<0.0001) but not in healthy subjects (broken line). The slopes of the regression lines are significantly different (p<0.01). (C) Frequencies of T cells producing interferon-γ (IFN-γ) in response to autologous Epstein–Barr virus (EBV) infected lymphoblastoid cell lines (LCLs) plotted against the proportion of CD8 T cells in the blood of EBV-seropositive healthy subjects (HC) (broken line) and EBV-seropositive patients with MS (continuous line). There is a significant positive correlation between the LCL-specific T cell frequency and the proportion of CD8 T cells in the healthy subjects (r=0.48, p<0.01) but not in the patients with MS (r=0.04, p=0.84). For a given proportion of CD8 T cells in the blood, the LCL-specific T cell frequency in MS patients is generally lower than in healthy subjects. This indicates that the decreased LCL-specific T cell frequency in MS is due to the CD8 T cell deficiency and a decreased proportion of LCL-specific T cells within the general CD8 T cell population. The slopes of the regression lines are significantly different (p<0.05). (D) Frequencies of T cells producing IFN-γ in response to autologous EBV-infected LCLs plotted against the age (in years) of 34 EBV-seropositive healthy subjects (HC) (broken line) and 34 EBV-seropositive patients with MS (continuous line). For HC, r=0.25 and p=0.15; for MS patients, r=0.17 and p=0.33. The y intercepts (elevations) of the regression lines are significantly different (p<0.01).
The frequency of PBMC producing IFN-γ in response to autologous LCL was available for 31 of the healthy subjects and 29 of the MS patients from our previous study.3 In the healthy subjects, there was a strong positive correlation between the LCL-specific T cell frequency and the proportion of CD8 T cells in the blood, but this correlation was lost in the MS patients (figure 1C). Figure 1C also shows that for a given percentage of CD8 T cells in the blood, the LCL-specific T cell frequency in MS patients was generally lower than in healthy subjects. This indicates that the decreased LCL-specific T cell frequency in MS is due to the CD8 T cell deficiency and a decreased proportion of LCL-specific T cells within the general CD8 T cell population. The LCL-specific T cell frequency was lower in MS patients than in healthy subjects at all ages but, in contrast to the proportion of CD8 T cells, did not decline with increasing age (figure 1D).
Discussion
The present study confirms the previously reported CD8 T cell deficiency in patients with MS4 and shows that it contributes to the decreased CD8 T cell response to EBV-infected B cells and worsens with age. Our finding that the CD8 T cell deficiency is most marked in SPMS and PPMS is consistent with previous studies showing that the decrease is most pronounced in progressive MS.4 What is remarkable about the CD8 T cell deficiency in MS is that normally when the EBV load is increased, as it is in the blood and brain of MS patients,1 the CD8 T cell frequency should increase and not decrease.5 This suggests that there is a fundamental defect in the ability of MS patients to make an appropriate CD8 T cell response to EBV, as we have previously hypothesised.1 2 We propose that this defect is genetically determined.
The absence of the normal correlation between the LCL-specific T cell frequency and the proportion of CD8 T cells in the blood of MS patients (figure 1C) indicates that, superimposed on the CD8 T cell deficiency, there is a deficiency of LCL-specific T cells within the general CD8 T cell population, which is most evident in SPMS. This may reflect T cell exhaustion, which occurs in virus-specific CD8 T cells during high-grade chronic viral infections. We found a marked aggravation of the CD8 T cell deficiency with increasing age in MS patients, particularly in PPMS. CD8 T cells normally decline slightly in number with increasing age although this was not statistically significant in our healthy subjects, probably because of the relatively low number of participants. The accelerated decline in the proportion of CD8 T cells with age in MS may partly result from deletion of EBV-specific T cells through exhaustion but may also be an intrinsic component of a genetic CD8 T cell deficiency. Accelerated age-related CD8 T cell deficiency and the consequent increasing EBV load might underlie the age-dependent accumulation of disability in MS.6
In conclusion, we have shown CD8 T cell deficiency contributes to the decreased CD8 T cell response to EBV-infected B cells and worsens with age in MS patients. These findings have implications for the prevention and treatment of MS by controlling EBV infection.1
Acknowledgments
We are grateful to Dr Kerryn Green, Dr Stefan Blum, Kaye Hooper and Bernie Gazzard for assistance in clinically assessing patients and in the collection of blood samples and to Dr Peter Baker for statistical advice. We apologise to those colleagues whose works were not cited owing to space limitation.
Footnotes
Funding This work was supported by project grants from Multiple Sclerosis Research Australia.
Competing interests None.
Ethics approval Royal Brisbane & Women's Hospital Human Research Ethics Committee and The University of Queensland Medical Research Ethics Committee.
Provenance and peer review Not commissioned; externally peer reviewed.