Article Text

Abstract
Facioscapulohumeral muscular dystrophy (FSHD) is caused by the size reduction of a polymorphic repeat array on 4q35. Probe p13E-11 recognises this chromosomal rearrangement and is generally used for diagnosis. However, diagnosis of FSHD is complicated by three factors. First, the probe cross hybridises to a highly homologous repeat array locus on chromosome 10q26. Second, although aBlnI polymorphism allows discrimination between the repeat units on chromosomes 4 and 10 and greatly facilitates FSHD diagnosis, the occurrence of translocations between chromosomes 4 and 10 further complicates accurate FSHD diagnosis. Third, the recent identification of deletions of p13E-11 in both control and FSHD populations is an additional complicating factor. Although pulsed field gel electrophoresis is very useful and sometimes necessary to detect these rearrangements, this technique is not operational in most FSHD diagnostic laboratories. Moreover, repeat arrays >200 kb are often difficult to detect and can falsely suggest a deletion of p13E-11. Therefore, we have developed an easy and reliable Southern blotting method to identify exchanges between 4 type and 10 type repeat arrays and deletions of p13E-11. ThisBglII-BlnI dosage test addresses all the above mentioned complicating factors and can be carried out in addition to the standard Southern blot analysis for FSHD diagnosis as performed in most laboratories. It will enhance the specificity and sensitivity of conventional FSHD diagnosis to the values obtained by PFGE based diagnosis of FSHD. Moreover, this study delimits the FSHD candidate gene region by mapping the 4;10 translocation breakpoint proximal to the polymorphicBlnI site in the first repeat unit.
- FSHD
- diagnosis
- dosage
- subtelomere
Statistics from Altmetric.com
Facioscapulohumeral muscular dystrophy (FSHD, MIM 158900) is the third most common inherited neuromuscular disorder, mainly characterised by a progressive weakness of the facial, shoulder girdle, and upper arm muscles.1 2 The major locus for this autosomal dominant disorder, FSHD1, maps to the subtelomeric region of the long arm of chromosome 4 (4q35).3 This subtelomere contains a polymorphic repeat array locus consisting of 3.3 kb repeat units (D4Z4). In unaffected subjects, the number of repeat units varies between 10 and 100 copies (giving array lengths of 35-300 kb). Patients carry one chromosome with 1-10 copies as the result of a deletion of an integral number of tandemly arrayed D4Z4 repeat units.4 5
Since D4Z4-like repeat units are dispersed over the genome, probe p13E-11 (D4F104S1) is generally used for molecular diagnosis of FSHD. This probe maps proximal to the D4Z4 repeat array and also recognises a highly homologous polymorphic repeat array of similar size on 10q26 and a constant Y specific fragment.4 6 7 The 10q26 homologous array has complicated FSHD diagnosis since a short repeat array on 10q26 is non-pathogenic. However, the identification of a chromosome 10 specific BlnI restriction site within each chromosome 10 derived repeat unit has facilitated FSHD diagnosis.8 Generally, for diagnostic analysis, genomic DNA is digested byEcoRI/HindIII and by EcoRI/BlnI double restriction. EcoRI andHindIII do not digest within the repeat array allowing size determination of the entire repeat array with probe p13E-11. After hybridisation, four fragments will be visualised, two from chromosome 4 and two from chromosome 10. OnEcoRI/BlnI double digestion, only the chromosome 4 type alleles will be resistant to digestion, while the chromosome 10 type alleles will be digested into 3.3 kb fragments.
Owing to the large size of the normal alleles, conventional Southern blot analysis will not show all fragments. Generally, in the FSHD population, all fragments except for the FSHD allele and 10 type alleles <50 kb comigrate at the top of the gel. In the past, we have developed a pulsed field gel electrophoresis (PFGE) based fragment analysis showing all four alleles. Recently, this analysis has led to the identification of a subtelomeric instability of the repeat arrays resulting in translocated repeat arrays on both chromosomes 4 and 10. In fact, 20% of the Dutch population carries a chromosome 4 type repeat array on chromosome 10 or, alternatively, a chromosome 10 type repeat array on chromosome 4.9 Hence, the discrimination between repeat arrays derived from chromosomes 4 and 10 is not absolutely specific and we have already identified FSHD patients with short 10 type or compound repeat arrays (that is, a mixture of 4 type and 10 type repeat units) on chromosome 4.9 10 A further complication for diagnosis is that we have identified both controls and patients in whom the p13E-11 region is deleted.10
PFGE analysis performed on aqueous DNA eliminates most of the above mentioned pitfalls for FSHD diagnosis. However, the experimental procedure is quite complex, relying on high quality high molecular weight DNA. Accordingly, most laboratories performing FSHD diagnosis have difficulties in implementing aqueous DNA in PFGE based technology. Therefore, we have developed an additional diagnostic test, theBglII-BlnI dosage test, to identify translocations of the repeat array or deletions of p13E-11 in patients with otherwise inconclusive clinical and molecular diagnostic results. Interestingly, this test further delineates the FSHD candidate region on 4q35 and suggests a common mechanism underlying the allele exchanges between 4q and 10q.
Methods
SUBJECTS
Controls (Nos 1-3, 5-9, figs 1-3) carrying standard D4Z4 repeat array distributions or translocated repeat arrays (that is, either chromosome 4 type repeat units on chromosome 10q or 10 type repeat units on chromosome 4q) respectively are from a random population. The isolated FSHD patient (No 4, figs 1-3) was ascertained through one of the Dutch neuromuscular centres. Family Rf100 carrying a deletion of p13E-11 has been described elsewhere.10
FSHD DIAGNOSIS BY STANDARD GEL ELECTROPHORESIS AND PFGE
Standard gel electrophoresis and PFGE analysis were performed as described previously9-11 with the exception that the DNA was transferred to a Nytran Plus Membrane (Schleicher & Schuell) instead of Hybond N+ (Amersham).
BGLII-BLNI DOSAGE TEST
Two micrograms of genomic DNA was digested overnight withBglII (Pharmacia) andBlnI (Amersham) according to the manufacturer’s instructions. DNA was separated on a 0.8% agarose gel (Seakem), denatured twice for 30 minutes in 0.6 mol/l NaCl/0.4 mol/l NaOH, and blotted for five hours in the same buffer onto a Nytran Plus Membrane (Schleicher & Schuell). After blotting, the membrane was hybridised with probe p13E-11 (D4F104S1)4 as previously described.10 The blots were washed with a final stringency of 1 × SSC/0.1% SDS at 65°C for 15 minutes to remove most cross hybridisation. After exposure on Phosphorimager screens, the relative intensity of the hybridising fragments was quantified by the ImageQuant program (Molecular Dynamics).
Intensities were corrected for the background. The standard ratio (Rs) between intensities of 4 and 10 specific signals was obtained from three subjects (1-3) carrying normal repeat array distributions (Rs=Σ (R1..R3)/3). This standard ratio was applied as a correction factor in all subsequent tests. In fig 3, for each sample the contribution of chromosome 4 specific signal to the sum of the chromosomes 4 (S4) and 10 (S10) signals was calculated by (S4/Rs)/(S4/Rs+S10)=S4/(S4+S10×Rs).
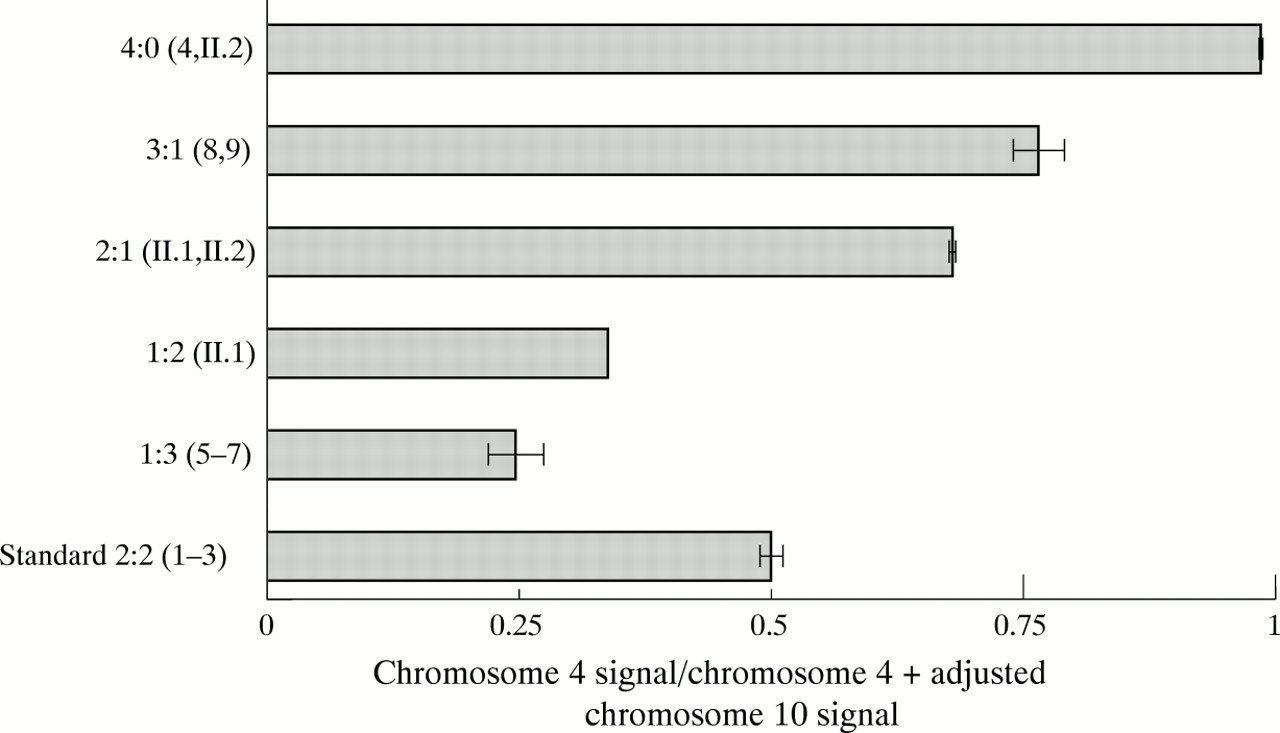
Relative intensities (chromosome 4 signal/chromosome 4 + adjusted chromosome 10 signal) of the different chromosome 4 and 10 constitutions in the same subjects as in fig 1. All the different samples produce the expected ratio as inferred from the PFGE analysis. Importantly, the confidence intervals (±2 SD) of the different ratios do not overlap.
Results
Fig 1A shows a regular Southern blot analysis as typically used for FSHD diagnosis. After digestion withEcoRI andEcoRI/BlnI double restriction, gels are run over 40 hours to facilitate the separation of fragments up to 50 kb. Lanes 1-3 and 5-9 on the left hand panel are healthy male controls selected from a random population. No 4 is a sporadic FSHD case. On the right hand panel, DNA of family Rf100 with a sporadic case of FSHD (II.1) has been loaded. This family has been described extensively elsewhere.10 The father, I.1, carries a deletion of p13-E11 on one of his chromosomes 4 which has been inherited by the sons, II.1 and II.2. Both I.1 and II.2 carry on this allele a D4Z4 repeat array >35 kb. However, in the affected son, II.1, the deletion is expanded in the D4Z4 repeat array reducing its size to 15 kb. Since this deletion also encompasses the p13E-11 region, this allele cannot be visualised by standard molecular FSHD diagnosis using probe p13E-11. The mother carries 4 type D4Z4 repeat arrays on both chromosomes 4 and 10 as inferred from PFGE and fluorescence in situ hybridisation (FISH) analysis.10 Therefore, all sons carry a 4 type repeat array on the maternally inherited chromosome 10.

(A) Conventional Southern blot based FSHD diagnosis. DNA from eight controls (Nos 1-3, 5-9), one isolated FSHD patient (No 4), and one family with a de novo case of FSHD (I.1, father; II.1, affected son, II.2 unaffected son, II.3 unaffected son; and I.2 mother) were digested with EcoRI/HindIII (H) or EcoRI/BlnI (B), respectively, and hybridised with probe p13E-11. The small de novo D4Z4 repeat array associated with FSHD in No 4 is marked with an arrow. The small 10 type repeat arrays in Nos 6 and 7 are marked with a small arrow while a cluster of 4 type D4Z4 repeat units derived from a compound repeat array in No 9 is marked with an asterisk. The size marker (lambda HindIII/PstI) is marked M. (B) PFGE based FSHD diagnosis of the same subjects as in (A). Note that the isolated FSHD patient No 4 carries four 4 type D4Z4 repeat arrays as inferred from the BlnI insensitivity of all four alleles. The same holds true for the mother I.2 of family Rf100 where two alleles comigrate at 65 kb. Nos 5, 6, and 7 carry three BlnI sensitive repeat arrays while No 8 carries only one. No 9 carries a compound repeat array consisting of both 4 type and 10 type units (asterisk). I.1, II.1, and II.2 carry a deletion of p13E-11. Owing to the large allele size, the upper allele No 1 is poorly visible (open circle). Marker lanes are indicated by an M.
As seen from this figure, the sporadic FSHD case (lane 4) can easily be recognised by the presence of a short D4Z4 repeat array. Subjects 6, 7, and 8 also carry a small fragment hybridising with probe p13E-11. However, since these fragments are sensitive toBlnI, they are probably residing on chromosome 10. Clearly, this test fails to identify the FSHD allele in the proband in family Rf100 (II.1) due to the deletion of p13E-11. Additional information cannot be obtained from this figure since large repeat array fragments comigrate on top of the gel.
Fig 1B shows the same samples run by PFGE. This gel clearly shows that PFGE is far more informative since it allows the discrimination of all four p13E-11 hybridising fragments. The sporadic FSHD case (lane 4) turns out to carry 4 type repeat arrays on both chromosomes 4 and 10 as judged from their BlnI insensitivity.
In fig 1A, if subjects 6, 7, and 8 were suspected FSHD cases, they would be regarded as unaffected based on theBlnI sensitivity of the small repeat array. However, fig 1B shows that subjects 6 and 7 carry three 10 type repeat arrays, one of which resides on chromosome 4. In contrast, subject 8 carries three 4 type alleles. Although these subjects are from a healthy control group and highly unlikely to be affected, suspected FSHD cases carrying such a complement of repeats could be given a false negative diagnosis by conventional Southern analysis. Therefore, further tests are required to confirm the chromosome 4 origin of the small fragment. This can be done by haplotype analysis or aNotI digest followed by a chromosome 4 specific hybridisation.10
Subjects 8 and 9 carry three BlnI resistant repeat arrays indicative of a 4 type repeat array on one of their chromosomes 10. Subject 8 carries a homogeneous translocated 4 type array. In subject 9, however, the presence of a smallBlnI resistant fragment (asterisk) indicates a hybrid repeat array consisting of several 4 type repeat units followed by a cluster of 10 type repeats. Such hybrid array constitutions have been reported recently.10 12
On the right hand panel of fig 1B, it is evident that the mother I.2 carries three BlnI resistant repeat arrays. In fact, detailed analysis has shown that the 65 kb fragment consists of two comigrating chromosome 4 alleles.10 Owing to the deletion of p13E-11, only three alleles are visible in I.1, II.1, and II.2 after hybridisation with probe p13E-11.
Although it is obvious that PFGE is highly informative, it is highly dependent on the quality of the aqueous DNA. Currently, only a limited number of laboratories successfully run PFGE based FSHD analysis. The quality of the aqueous DNA is particularly important for fragments >200 kb. This is also shown in fig 1B where in subject 1 only three of the four alleles are recognised. This may erroneously suggest a deletion of p13E-11.
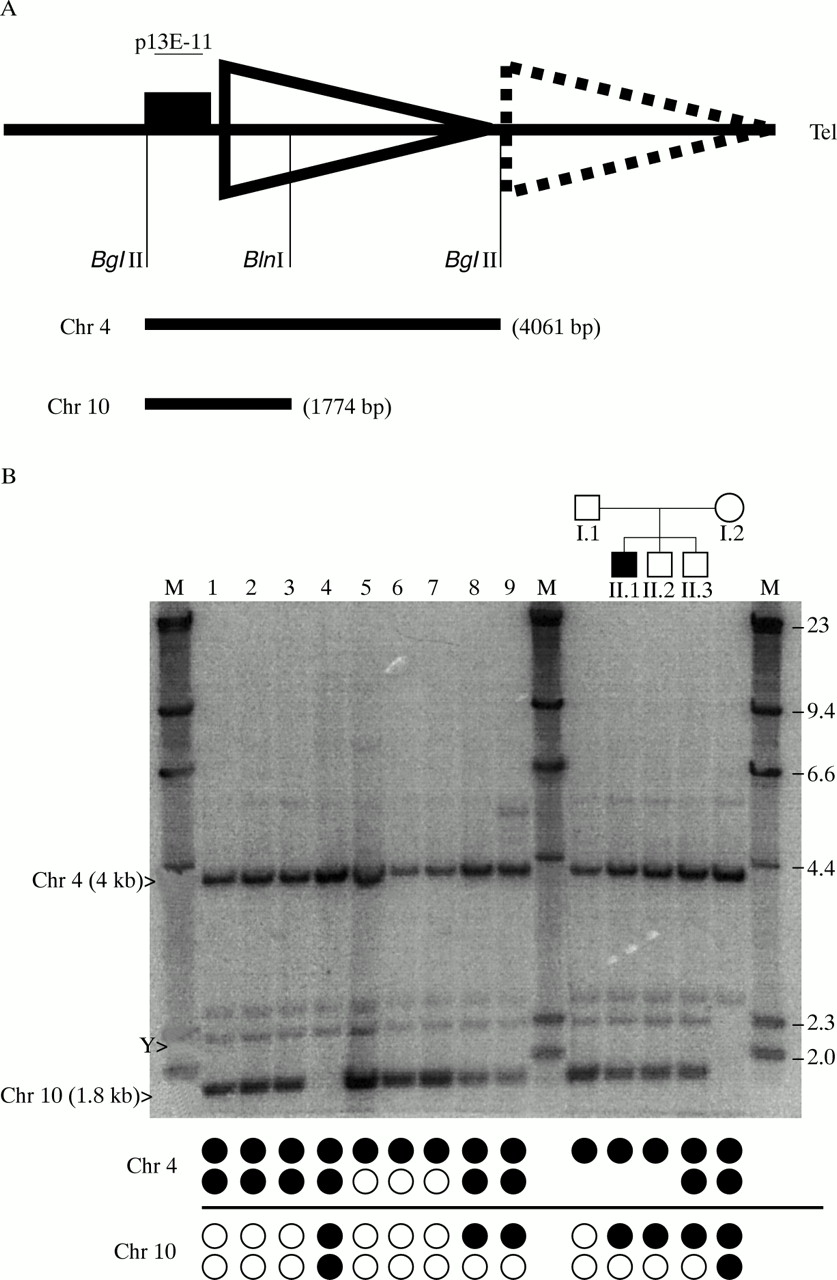
To avoid the false identification of carriers of deletions of p13E-11, and to design a simple test to identify those carrying translocated alleles, we developed a novel additional Southern blot test, theBglII-BlnI dosage test, that can be run on aqueous DNA in any laboratory. The test uses the BlnI polymorphism in the first D4Z4 repeat unit and BglII instead ofEcoRI to obtain a small sized fragment (fig2A). Double digestion with BglII andBlnI will release a chromosome 4 derived fragment of 4061 bp after hybridisation with p13E-11. Chromosome 10 derived fragments are, owing to the BlnI site, only 1774 bp in size. Fig 2B shows DNA of the same subjects as in figs 1A and B after digestion with BglII andBlnI and hybridisation with p13E-11. In subjects without translocated alleles and deletions of p13E-11 (Nos 1, 2, and 3), the ratios between the signal intensities from the chromosome 4 (4 kb) and 10 (1.8 kb) fragments should be 2:2. In those carrying one or three 4 type repeat arrays, the 4:10 ratios should be 1:3 and 3:1, respectively. Indeed, these different intensities of the 4 derived and 10 derived signals are clear even by visual inspection. Subjects 5-9, carrying three 4 type or 10 type alleles, respectively, show skewed intensity ratios. Subject 4, carrying only 4 type repeat arrays shows no hybridisation at the 10 type repeat unit derived fragment length (1.8 kb). These observations are confirmed by computer aided intensity measurements using the ImageQuant program as shown in table 1. Here, the calculated 4:10 ratios perfectly reflect the expected values based upon PFGE.

{kind=link}
{kind=link}
{kind=link}
(A) Schematic overview of the BglII-BlnI dosage test. DNA is digested with BglII and BlnI releasing the p13E-11 region including the first D4Z4 repeat array. Owing to the presence of a polymorphic BlnI site in the chromosome 10 type repeat unit but not in the 4 type repeat unit, this restriction will release a 4061 bp fragment from chromosome 4 and a 1774 bp fragment from chromosome 10 indicated by the bars underneath. The region hybridising with p13E-11 is indicated by a filled box. (B) Southern blot of a BglII-BlnI dosage test of the same subjects as in fig 1. The chromosome 4 (4061 bp) and chromosome 10 (1774 bp) derived fragments are indicated with their respective chromosome numbers. The cross hybridising chromosome Y fragment is indicated by a Y. Signal intensities from the fragments of both chromosomes can be compared to evaluate the presence of translocated D4Z4 alleles or deletions of the region spanning the probe p13E-11. Underneath the lanes, the repeat array constitutions of the different alleles can be seen. Filled circles represent 4 type arrays, open circles are 10 type arrays on both chromosomes.
Expected and observed ratios between the chromosome 4 and 10 derived signal intensities after correction for the background. The first row shows the same subjects as figs 1-3. The second row lists the different repeat array constitutions as inferred from PFGE (fig 1B). The third row indicates the expected 4:10 ratios while the fourth row shows the 4:10 ratios obtained by the BglII-BlnI dosage test
The BglII/BlnI dosage test also allows the identification of complex rearrangements such as deletions of the p13E-11 region, as seen on the right hand panel of fig 2B. As mentioned, the father carries a deletion of the p13E-11 region on one of his chromosomes 4 and the mother only carries 4 type repeat arrays. Therefore, the 4:10 ratio in the father should be 1:2 and in the mother 4:0. In the two sons (II.1 and II.2) carrying the deletion of p13E-11 and a 4 type repeat array on the maternally inherited chromosome 10, the ratio should be 2:1, and in the son II.3 a ratio of 3:1 is expected. Indeed, as seen in table 1, the expected ratios match the ratios obtained by the ImageQuant program.
Apart from the chromosome 4 and 10 derived fragments, some cross hybridising fragments are also visible that do not interfere with this test. One of the fragments is derived from the Y chromosome as indicated by the absence of this fragment in the only female (I.2).
Fig 3 shows the contribution of the 4 type p13E-11 derived signal to the total (4 type and 10 type) p13E-11 derived signal intensities for different allele constitutions in a bar chart. All ratios closely match the expected values. Importantly, the confidence intervals (± 2 SD) of all different repeat constitutions do not overlap.
So far, we have tested 204 chromosome 4 and 10 alleles of patients and controls, of which 35 were translocated to the non-homologous chromosome and three were deleted for the p13E-11 region as inferred from our PFGE data. Of these 35 translocated alleles, four repeat arrays were composed of clusters of 4 type and 10 type repeat units. If translocations between chromosomes 4 and 10 had occurred distal to the first polymorphic BlnI site, this would not result in a dosage difference of the 4 derived and 10 derivedBglII fragments. However, in all cases theBglII/BlnI dosage test is consistent with 4;10 translocations occurring proximal to the first polymorphic BlnI site.
Discussion
FSHD diagnosis relies on the detection of a D4Z4 repeat array <35 kb (<10 units). For this analysis, the discrimination between 4 type and 10 type repeats is essential and is based on a 10 type repeat unit specific BlnI site. However, our PFGE data suggest a dynamic recurrent instability of the subtelomeric D4Z4 repeat array involving both chromosomes 4 and 10. This instability results in translocated 4 type repeat arrays on chromosome 10 and vice versa in 20% of the Dutch population. Interestingly, short 10 type repeat arrays or short compound repeat arrays consisting of both types of repeat units residing on chromosome 4 also cause FSHD. However, similar small repeat arrays on chromosome 10 are non-pathogenic, irrespective of the type of repeat.9 Thus, in suspected FSHD patients carrying three 10 type repeat arrays, a short 10 type repeat may reside on chromosome 4 resulting in FSHD.
PFGE based FSHD diagnosis has the advantage that all four alleles are visualised. However, the success relies heavily on the quality of the aqueous DNA and often large alleles exceeding 200 kb are difficult to detect. Also, many diagnostic laboratories do not have the facilities, nor the expertise, successfully to carry out such PFGE based FSHD diagnosis. Therefore, we have developed an additional test, theBglII-BlnI dosage test, which easily recognises deletions of p13E-11 or translocated alleles and which can be used in case of inconclusive results.
The BglII-BlnI dosage test uses the BlnI polymorphism within the first repeat unit adjacent to p13E-11 to differentiate between 4 type and 10 type repeat units. Digestion byBglII releases a fragment of 4061 bp carrying p13E-11 and the first D4Z4 repeat unit. Subsequent digestion with BlnI will reduce the size of the 10 type fragments to 1774 bp, while the 4 type fragments remain undigested. After standard agarose gel electrophoresis and hybridisation with p13E-11, the relative intensities of the two fragments can easily be scored by visual inspection or quantitative analysis programs. Normally the 4:10 ratio should be 1 (2:2), subjects carrying three 4 type repeat arrays have a ratio of 3 (3:1), while subjects carrying three 10 type repeat arrays have a ratio of 0.33 (1:3). The data we present here show the integrity of the test. Even complex rearrangements including translocations of the repeat arrays and deletions of p13E-11 are recognised by the skewed ratios. Although the number of observations in each group of rearrangements was small, it is still important to observe (fig 3) that the confidence intervals for the estimated dosage ratio in each of the different groups never overlap, while they almost always contain the theoretically expected “true” ratio (except for the group with an expected ratio of 1, where background may easily lead to small deviations). The observed accuracy of the test is probably mainly because of the quantitative intensity analysis within one sample and not between samples, avoiding quality differences in the DNA. Although quantification of the ratios by a densitometer is desirable, common rearrangements such as translocations may readily be scored by visual inspection. Therefore, using theBglII-BlnI dosage test, large populations can easily be tested for the presence of translocated alleles. Moreover, this test can be used in addition to the PFGE based FSHD analysis to prevent false identification of deletions of p13E-11 owing to the difficult visualisation of large fragment sizes (>200 kb).
The BglII-BlnI dosage test will only fall short in subjects who carry complex rearrangements in which a 4 type repeat resides on chromosome 10 and a 10 type repeat on chromosome 4, and in subjects carrying a deletion of p13E-11 and identical repeat type alleles on the other three chromosomes. The PFGE based diagnosis of FSHD usingBlnI to discriminate between 4 type and 10 type alleles has a sensitivity of 96.5% and a specificity of >99%. In contrast, the sensitivity of the same diagnosis using conventional linear gel electrophoresis is 92% with a specificity of 99%.13 Adding theBglII/BlnI dosage test to the conventional diagnosis will raise the sensitivity and specificity to the values reached by PFGE.
PFGE has already indicated that in most cases the entire repeat array was translocated to the non-homologous chromosome. Only in a subset of the translocations did the repeat arrays consist of hybrid clusters of 4 type and 10 type repeat units.10 However, formerly, we could not exclude a translocation within the repeat array itself. An interesting implication from the robustness of this test is that translocations between chromosomes 4 and 10 must have occurred proximal to the polymorphic BlnI site within the first repeat unit. So far, we have analysed a selected set of 51 patients and controls by theBglII-BlnI dosage test. Twenty eight of them carried translocated alleles of which four consisted of both types of repeat. None of the 4:10 ratios observed differs from the expected ratio. Even in the four subjects carrying compound translocated alleles (that is, arrays consisting of alternating 4 type and 10 type repeat units), the translocation had taken place proximal to the first polymorphicBlnI site. Thus, it is likely that in all cases the entire repeat is translocated to the non-homologous chromosome. Since the homology between both chromosomes only extends 40 kb proximal to the repeat array, this implies either a common origin of 4q and 10q translocated alleles (founder effect) or, in the case of recurrent translocations, a recombination hotspot within this 40 kb segment. The exact localisation of this translocation breakpoint proximal to the repeat arrays has implications for the localisation of the putative FSHD gene. It is generally hypothesised that FSHD is caused by a position effect in which deletions of the D4Z4 repeat influence the transcription of genes nearby. The chromosome 4 specificity of the disease implies that the FSHD gene must be located proximal to the translocation breakpoint. Therefore, precise mapping of this translocation breakpoint will refine the FSHD candidate gene region.
Interestingly, in yeast it has been shown that recombination hotspots colocalise with open chromatin domains, often promoter or coding sequences. In vertebrates, a similar mechanism may play a role in recombination.14 Thus, a putative recombination hotspot proximal to the D4Z4 array may indicate a new FSHD candidate gene locus.
Acknowledgments
This study was funded by the Prinses Beatrix Fonds, The Dutch Organization for Scientific Research (NWO), The Muscular Dystrophy Association (USA), The Dutch FSHD Foundation, The FSH Society, and the Association Française contre les Myopathies (AFM).