Article Text

Abstract
Visual perceptual disorders are often presented as a disparate group of neurological deficits with little consideration given to the wide range of visual symptoms found in psychiatric and neurodevelopmental disease. Here, the authors attempt a functional anatomical classification of all disorders linked to visual perception, whatever the clinical context in which they arise, including those disorders that bridge vision, emotion, memory, language and action. Guided by clinical and neuroimaging evidence, visual perceptual disorders are classified by the functional anatomical networks likely to be involved and the class of underlying dysfunction, whether topological (a localised deficit or region of hyperfunction) or hodological (a disconnection or hyperconnection). The wider perspective forces us to consider what visual functions underlie a range of symptoms sidelined by previous classificatory schemes and helps generate novel hypotheses for further research in the area.
- Hallucinations
- neuroanatomy
- neuropsychiatry
- vision
Statistics from Altmetric.com
Introduction
If visual perception is the day-to-day experience of ‘seeing’—or put another way, the conscious, visual experience of the world we have when we open our eyes—a visual perceptual disorder might be considered a deviation from this experience. Current classificatory schemes focus on disorders traditionally considered neurological (ie, based on static visual deficits typically related to brain lesions), relating each disorder to a specific brain region or its connections.1 2 Visual symptoms more likely to be found in psychiatric or neurodevelopmental disorders such as visual hallucinations, illusions and visual perceptual distortions, are typically only mentioned in passing or not at all in such schemes. This omission is understandable given that these types of symptoms are often transient and thus difficult to study directly, and may occur without an obvious associated brain lesion. Here we attempt to incorporate both ‘neurological’ and ‘psychiatric’ disorders into a single classificatory scheme using an anatomical network approach. Our aim is to provide an anatomically structured overview of visual perceptual dysfunction across all clinical contexts that we hope will provoke further research.
A note on terminology
The scope of our classification is set by the way we have interpreted the three constituent terms of ‘visual perceptual disorder.’
Visual
The disorders classified here are visual in the sense that they are selective for vision without equivalent changes in other sensory modalities. For example, visual hypoemotionality (a loss of emotional tone for visual stimuli) implies that, in some cases at least, emotions may be preserved for auditory and other sensory stimuli.
Perceptual
In deviating from normal ‘seeing', any cause of visual impairment meets the definition of a ‘visual perceptual disorder' outlined above, for example, a blurring of vision through loss of acuity. To limit the scope of the review, we have focused on symptoms attributable to dysfunction within cortico-cortical or cortico-subcortical networks including those that bridge visual perception with memory, attention, emotion and action. We make a distinction between visual perception and visual imagery, as the two differ in terms of their underlying neurobiology.3 Visual percepts are seen in the external world and are outside volitional control (one cannot change the percept by force of will), whereas visual imagery is seen in the mind's eye and is under volitional control. We consider visual hallucinations and illusions as varieties of visual perceptual experience, as they are closely related to visual perception in terms of underlying brain activity.3 Phenomena with elements of both perception and imagery are more difficult to classify. For example, experiences referred to as pseudohallucinations or incomplete hallucinations (see Blom4 for other uses of these terms) are seen in the mind's eye but are outside volitional control. We have highlighted such ambiguous experiences in the classification and, where appropriate, presented both perceptual and imagery viewpoints.
Disorder
We use the term disorder for continuity with previous clinical literature, although much of the classification relates to perceptual symptoms that do not imply a specific disease or pathology.
Hodotopic framework
We have classified visual perceptual disorders using the general anatomical framework we set out in 2005 as an update of classical associationist and neo-associationist accounts of higher cognitive function and dysfunction.5 Recent advances in functional neuroanatomy, using techniques such as functional MRI (fMRI) and diffusion tensor tractography (DT-tractography), provide complementary views of the anatomy of higher function and the pathological mechanisms that generate cognitive and behavioural symptoms. One view, termed topological (from ‘topos’=place), relates higher function and dysfunction to discrete cortical locations. The other, termed hodological (from ‘hodos’=path), relates higher function and dysfunction to connections between locations. Both views are combined in a generic neuroanatomical model applicable to all higher functions and dysfunctions—the hodotopic framework.5 From the perspective of the hodotopic framework, each higher function relates to a specific network of cortical locations and white-matter connections. Topological disorders are localised within a cortical area and result in a loss (deficit) or an increase in function (hyperfunction). Hodological disorders relate to connections between areas, including those in which function in one brain region is altered by changes in another, spatially remote, region. We use von Monakow's term ‘diaschisis’ to refer to such remote effects6; however, the term is not intended here to carry its original implication of transient neurological shock. Connections between areas can be decreased (disconnection) or increased (hyperconnection) in hodological disorders. Deficit and disconnection disorders of visual perception are found in stroke, intracranial compressing masses, neurosurgical procedures, demyelinating disorders, neurodevelopmental conditions (eg, autism), neurodegenerative disease, schizophrenia and depression. Hyperfunction and hyperconnection disorders of visual perception are found in ophthalmic disease, neurodegenerative disease, Parkinson's disease, psychosis, epilepsy and as normal phenomena in relation to sleep or in the context of psychedelic drugs, alcohol or medication side-effects.
Classification method
We have allocated each visual perceptual symptom to a visual network and one of four classes of dysfunction (topological versus hodological; hyper- versus hypo- function/connection). For some symptoms, functional imaging, DT-tractography or lesion evidence is available to help identify the appropriate location in the table (eg, a localised increase in activity within a specific brain region or a decrease in connectivity within a specific tract). However, for many, we have been forced to make an educated guess through: (1) analogies to related symptoms for which evidence is available and (2) a neurophenomenological approach.3 7 This approach is derived from the body of research into the neural correlates of consciousness (NCC) which suggests that some, as yet undefined, aspect of activity within a specialised cortical region (resulting in an increase in imaging signal8) contributes directly to visual perceptual experience (eg, the percept of a face is associated with increased activity within face-specialised cortex9). Where evidence is available, this principle seems also to apply to hallucinatory, illusory and synaesthetic percepts.3 Thus, the content of visual perceptual experience (whether an object, a face, motion, etc) localises activity to a particular brain region or network, irrespective of whether the experience is a normal percept, illusion, hallucination, etc.
A taxonomy of disordered visual perception
Table 1 and figure 1 outline a tentative classification of visual perceptual disorders.
Disorders of visual perception
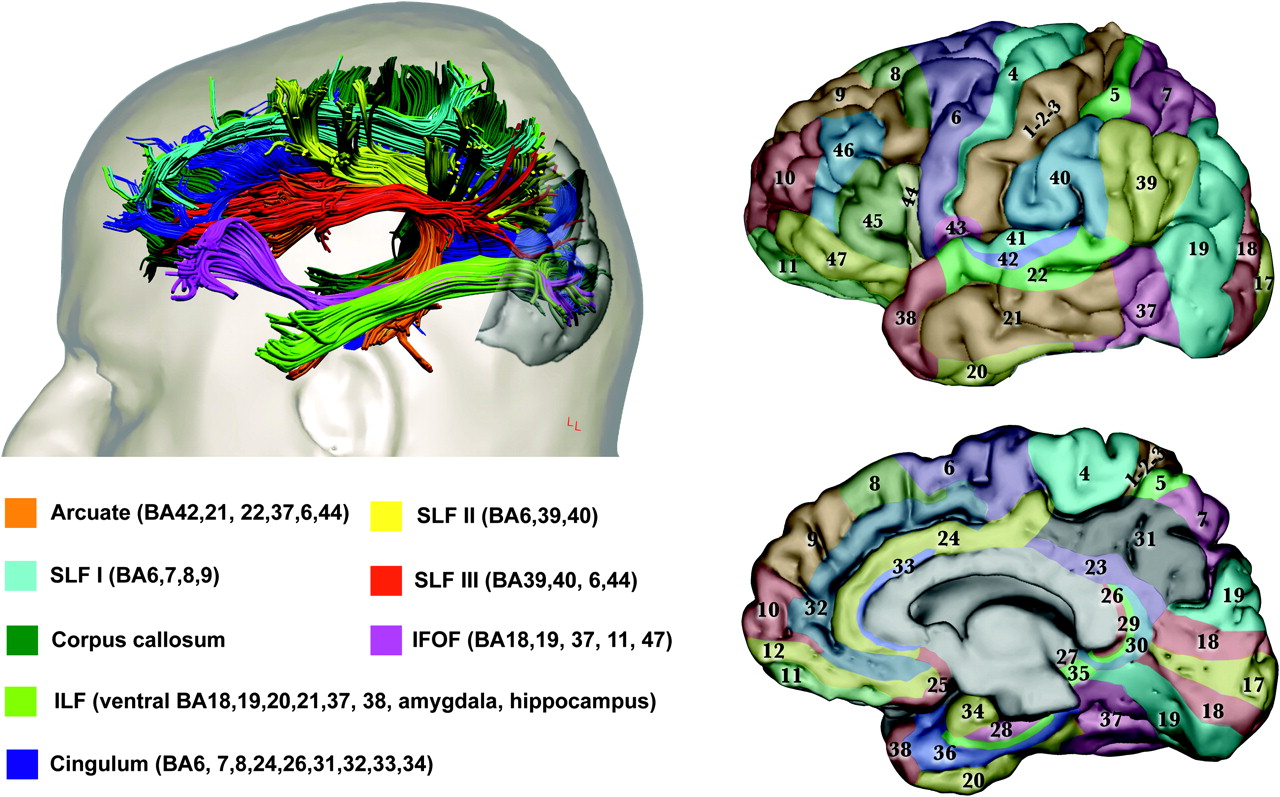
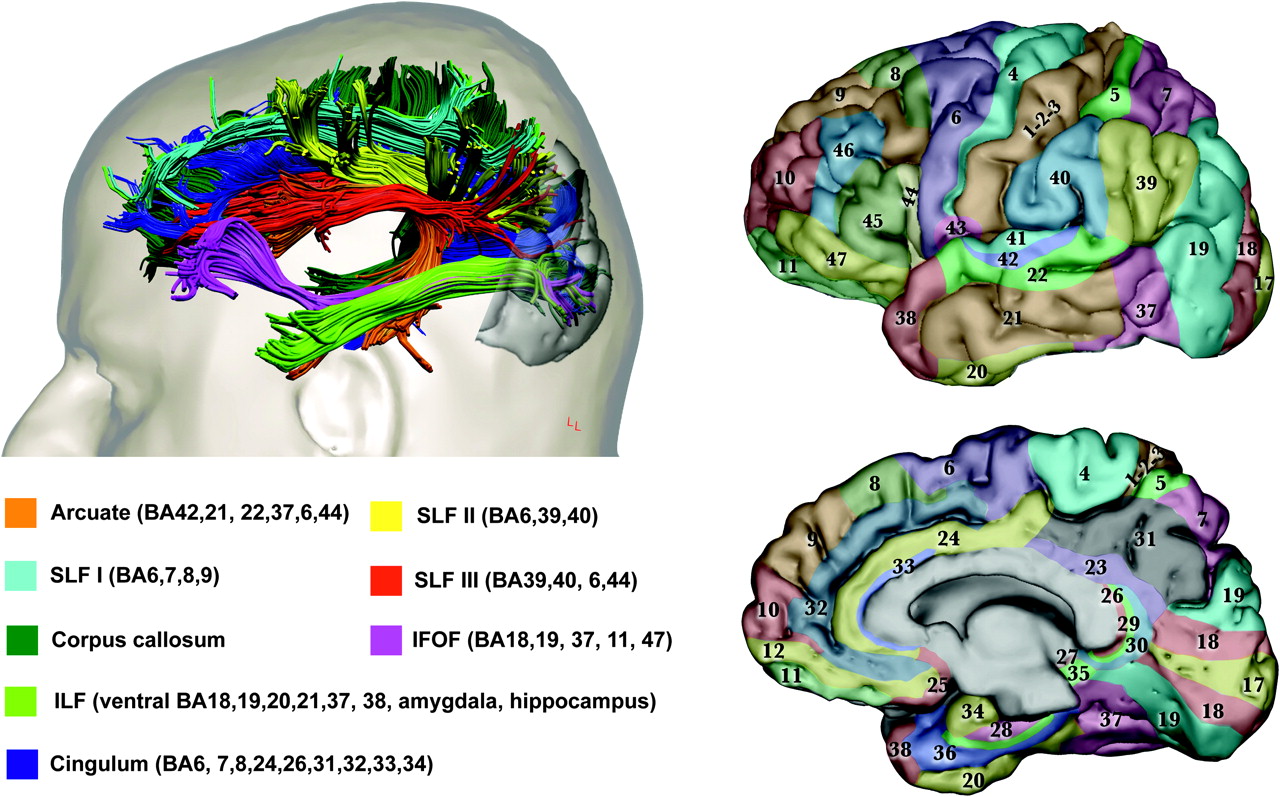{kind=link}
Anatomy of visual networks. The upper left panel shows the major associative tracts with connection to the occipital lobe in the human brain. The Brodmann areas corresponding to the cortical projections of each fibre tract are given in the lower left panel. The right panels show Brodmann areas colour-coded on lateral (upper) and medial (lower) views of the brain. Images modified from Catani and Thiebaut de Schotten.10 The primary visual cortex (V1, area 17) is located on the polar and medial surface of the occipital lobe. V2 (within BA18) is adjacent to V1 and is divided on the medial occipital surface into a dorsal portion and a ventral portion. V3 (within BA18) is adjacent to V2 and is similarly divided. The occipital lobe, lateral and ventral occipitotemporal cortex, limbic cortex and the parietal lobe contain regions specialised for different visual perceptual attributes, including: familiar faces, colours, extended landscapes, objects, text and visual textures (ventral occipito-temporal cortex) and visual motion, body parts, face features, intermediate-level object processing, and eye and mouth movements (posterior-lateral cortex).7 The predominant pathway from the eye to V1 relays through the lateral geniculate nucleus (LGN), although a clinically important pathway passes directly from the LGN or pulvinar to motion-specialised cortex (V5 within BA 19/37) on the lateral occipital surface.11 The detailed anatomy of fibres within and between occipital regions and from the occipital lobe to the pulvinar has yet to be characterised in man. Chains of U-shaped fibres and parallel long association pathways passing to the parietal lobe form a cortical network (dorsal stream)12 thought to underlie spatial aspects of vision, while equivalent pathways to the temporal lobe form a cortical network (ventral stream) thought to underlie object vision, colours, faces and visual language. The inferior longitudinal fasciculus (ILF) carries reciprocal connections between the occipital and medial/lateral anterior temporal lobes related to visual emotion and visual memory.13 The inferior fronto-occipital fasciculus (IFOF) connects territories in the occipital lobe and ventral frontal lobe.12 Connections between the visual parietal lobe and frontal cortex are carried in the superior longitudinal fasciculus (SLF), while those between occipital and limbic cortex are carried in the cingulum and ILF. The SLF and IFOF are likely to relate to functions such as visual attention, visual working memory, visual praxis, visual-vestibular function and eye movement control. Cortico-cortical visual connections between hemispheres pass predominantly through the splenium of the corpus callosum.
Topological disorders of visual perception
Deficits
Prototypical disorders of this nature are characterised by the selective loss of a specific visual perceptual function—for example, achromatopsia14 (selective loss of colour vision following lesions of colour-specialised cortex) and akinetopsia15 (selective loss of motion vision following lesions of lateral occipital motion-specialised cortex; the motion deficit is not absolute with a relative sparing of the perception of slow movement16).
Lesions of V1 result in a scotoma restricted to a single hemifield (a hemianopia if the lesion involves the entire extent of V1 in one hemisphere). Lesions of ventral V2/V3 (within subcalcarine BA18) are associated with upper quadrantic field defects, while those of dorsal V2/V3 (supracalcarine BA18) are associated with inferior quadrantic field detects.17 Scotomata in the context of migraine aura relate to regions of cortical spreading depression-like activity in early visual areas (eg, V1, V2 and V318).
Face perception deficits (the prosopagnosias) have a variety of subtypes likely to reflect the involvement of different functional modules within the face-processing system, although disconnection mechanisms may also be involved.19
An inability to recognise familiar places (environmental agnosia) is associated with medial temporal lobe lesions20 and can be considered a topological deficit of cortex specialised for the visual environment (parahippocampal place area21) or a hodological disconnection of visual and memory representations of places20 within the hippocampal-retrosplenial network (ventral cingulum).
Size and object distortions are referred to collectively as metamorphopsias. Objects appearing larger or smaller (macropsia and micropsia respectively) have been attributed to a dysfunction of object constancy within the temporal lobe (the integration of object distance with retinal visual angle to estimate size), although it is unclear whether this is best conceived as a topological dysfunction (ie, a deficit or hyperfunction of a cortical region specialised for constancy) or a hodological dysfunction (ie, a disconnection or hyperconnection of relevant specialised modules). The same mechanism may be responsible for objects appearing nearer or further away (pelopsia and teleopsia respectively), although such symptoms can also be interpreted as disorders of depth perception (and are also referred to as enhanced stereopsis4). Lesions of lateral occipito-temporal cortex may cause micropsia in one hemifield22 or bilaterally,23 although as object constancy may be preserved,23 such cases may relate to a deficit of object processing rather than constancy.
Apperceptive agnosia and cortical blindness describe diffuse visual loss affecting many visual domains. They occur in the context of extensive occipital lesions and are likely to involve both topological and hodological dysfunction. Several subtypes of disordered form perception have been described,24 each likely to depend on the form perception submodule(s) involved in the lesion. Scieropia refers to darkening of vision and follows diffuse occipital damage (see Wapner et al25 for an example: a patient whose vision was dimmed as if seeing everything at twilight). The same symptom occurring without other visual impairments (scierneuropsia) was described in the psychoanalytic literature26 and is likely to be an example of derealisation (see below).
The Riddoch syndrome is a variant of cortical blindness in which motion-specialised cortex (V5) remains intact and is activated through direct connections from the LGN and/or pulvinar.11 Subjects are able to consciously perceive and discriminate visual motion in their otherwise blind visual field when stimulated with fast motion,11 while under other stimulus conditions report visual percepts without the ability to discriminate (gnosanopsia) or have intact discrimination ability without visual percepts (blindsight or agnosopsia).11
In visuo-spatial neglect, patients have a deficiency of orienting attention to one hemifield (usually left) and (may) fail to perceive stimuli in this region when rival stimuli are present in the non-neglected hemifield (extinction). The disorder has been interpreted as topological (related to the cortical modules involved in visuospatial processing and attention—see Mort et al27 for a review of regions implicated) but hodological mechanisms have also been suggested.28 Simultanagnosia (an inability to sustain attention across different locations in the visual field) leads to objects appearing fragmented or disappearing from view29 with shrinkage of the visual attentional field.30 The disorder is likely to relate to parietal lobe specialisations for spatial visual attention, although it is unclear whether the disorder is best conceived as topological or hodological. Rapid fading of vision found with occipito-parietal lesions31 is likely to be caused by the same mechanism as simultanagnosia.
Selective lesions of the amygdala are associated with perceptual deficits of emotional facial expression32 with similar deficits described in autism spectrum disorder and psychopathy.
Visual imagery may be selectively impaired (ie, with intact auditory, olfactory, gustatory, etc imagery; see Basso et al33 for a review of case reports) with a relative preservation of visual perception. Loss of visual imagery may also be accompanied by a specific loss of visual dreaming (visual anoneria34) or a more general loss of dreams in all sensory modalities (Charcot–Wilbrand syndrome35). Such disorders can be interpreted as topological deficits of visual imagery generation or hodological disconnections within visual memory networks.
Hyperfunction
Topological disorders of hyperfunction are related to pathophysiological mechanisms such as local irritation (eg, in epilepsy or at the margins of a cortical lesion) and deafferentation caused by eye disease (Charles Bonnet Syndrome) or a visual pathway lesion; the mechanism by which loss of visual input results in increased cortical activity is unclear but may relate to changes in thalamocortical circuitry.36
Hallucinations linked to V1 and surrounding areas (V2/V3) are of simple featureless forms, colours and television-like static (visual snow). These phenomena are termed phosphenes or photopsias by some authors, although they are perhaps better referred to as simple hallucinations, given that they differ from other types of visual hallucination only in the location of cortex involved.36 In migraine aura, zigzag lines and patterns (teichopsia37) relate to the margins of a region of cortical spreading depression-like activity in early visual cortex.18
Colour, object and landscape hallucinations are each linked to their respective region of cortical specialisation,36 38 39 and the same is likely to be the case for motion hallucinations and passage hallucinations (visual hallucinations of persons or animals that pass sideways out of the visual field). Increased vividness of an attribute (eg, increased vividness of colours: hyperchromatopsia40) is thought to relate to increased activity within the relevant specialised region (eg, colour-specialised cortex for hyperchromatopsia). The origin of lilliputian hallucinations (small figures, often in costume) is unclear but has been classified here as related to object specialisations. The reduced size of such hallucinations may relate to visual constancy mechanisms.40
Text or letter-string hallucinations lacking semantic content are linked to the cortical area specialised for word forms, while those with semantic content (visual command hallucinations and visual verbal hallucinations4) are linked to higher-level representations of visual language.41
Illusions of distorted faces (prosopometamorphopsia) relate to activity on the lateral convexity of the occipital lobe,42 a region specialised for face features (the occipital face area), and the same region is likely to account for hallucinations of distorted faces. Hallucinations of normal (undistorted) faces relate to activity within ventral occipito-temporal cortex (fusiform face area),38 and the same region is likely to account for facial intermetamorphosis (a change in the visually perceived identity of a face).
A range of symptoms collectively termed palinopsia40 43 have been linked to dysfunction within parietal-lobe coordinate systems.44 They include: (1) polyopia (perceiving multiple copies of the same object often arranged in rows and columns; if the number of copies is large (eg, >100) it is referred to as entomopia45); (2) visual perseveration (an object remaining fixed in retinal coordinates with eye movements); (3) delayed palinopsia (an object returning to field of view); and (4) illusory visual spread (the spread of a pattern from an object to its surroundings). Parietal lobe regions are also likely to underlie the trailing phenomenon (a variety of visual perseveration in which a series of discontinuous stationary images trail behind a moving object) and positive afterimages (afterimages in which objects are seen in their true as opposed to complementary colours).
Patients with Anton's syndrome (visual anosognosia/denial of blindness in the context of cortical blindness) report visual experiences that are traditionally interpreted as confabulations (false memories or false reports of visual perceptual experience) or visual imagery experiences.46 These symptoms are likely to be the result of topological hyperfunction within visual memory systems. Flashbulb memories (detail-perfect memories of events that at the time of witnessing evoked a high level of surprise, consequentiality and/or emotional arousal47) and memory hallucinations (fantastic false memories conjured up retrospectively4) may relate to a similar hyperfunction of visual memory systems (and/or visual emotion systems in the case of flashbulb memories). Reperceptive hallucinations involve the replay of memory fragments (also termed experiential hallucinations4); however, it is unclear whether such experiences are visual imagery or visual percepts.48 If visual imagery, they are likely to involve hyperfunction of memory systems; if visual percepts, they are likely to involve hyperconnection between visual memory networks and occipital cortex.
Autoscopic phenomena describe a range of experiences in which the self is duplicated in external space. In autoscopy, the duplicate self is seen in the external world. In an out-of-body experience (OBE) the physical body is seen from the perspective of the duplicated external self. In heautoscopy the hallucinated ‘double’ is identified as oneself, despite the lack of an exact physical resemblance, and the subjective viewpoint may change between the double and physical self in rapid alternation. These phenomena are thought to relate to the disintegration of visual, proprioceptive, tactile and vestibular modalities, and have been linked to transient dysfunction in the region of the temporoparietal junction (TPJ).49 Whether this dysfunction is better conceived as topological hyperfunction of one or more of the sensory modalities involved or a hodological connectivity increase between specialised regions is unclear. Visual extracampine hallucinations (hallucinations perceived in impossible locations outside the field of view) could be considered related phenomena in that they involve an alteration in the reconstruction of visual space and body schema. Other disorders likely to be related to hyperfunction or hyperconnection within the TPJ and related regions include: (1) the oculogravic illusion (an illusory displacement of objects co-occurring with a change in gravity as in a diving airplane50); (2) the oculogyral illusion (the apparent movement of a stationary light in a dark room during fixation following a period of rotation50); (3) visual hallucinations occurring in association with caloric stimulation51; (4) Zingerle's automatosis in which visual hallucinations occur during transient motor and vestibular dysfunction3 (eg, compulsive posturing, torticollis, choreo-athetosis); (5) the transient intensification of visual patterns and text, often in association with neuroleptic induced oculogyric crises (paroxysmal perceptual alteration52).
Hodological disorders of visual perception
Disconnection
Disconnection of cortical visual networks from subcortical inputs as a result of visual pathway lesions or eye disease is associated with simple and complex visual hallucinations and visual illusions3 7 (the Charles Bonnet Syndrome—the change in cortical function secondary to eye disease might be considered a form of diaschisis). Brainstem lesions are associated with peduncular hallucinations (complex hallucinations associated with eye movement disorders and changes in arousal or sleep53).
Depth vision (visual space) involves a number of dissociable54 functions, some related to perceptual aspects of depth (eg, stereopsis (binocular vision) and object constancy), others related to action aspects of depth (eg, reaching behaviour). One aspect of depth vision (stereopsis) is lost in the central visual field (but not the visual periphery) following splenial section.55
In cinematographic vision, moving objects appear as a series of static freeze frames at different spatial locations.4 The disorder may relate to the disconnection of motion specialised cortex and spatial coordinate frames in the parietal lobe.
Associative object agnosia (loss of object meaning with intact object perception) is thought to relate to the disconnection of visual and semantic regions (see Riddoch and Humphreys24 for a review). Specific deficits of colour or object naming (colour anomia, object anomia) are thought to relate to the disconnection of visual and language areas, while optic aphasia (an inability to name objects by sight with preserved ability to name by touch or from description) is thought to follow disconnection of left-hemisphere verbal semantic and right-hemisphere visual semantic representations.56
Pure alexia (a deficit of reading words with preserved writing and letter recognition) and left hemialexia (a reading deficit restricted to the left hemifield) have been interpreted as hodological dysfunctions (a disconnection of a left-hemisphere region specialised for word forms from visual inputs) and topological dysfunctions (a deficit of the word form area) (see Epelbaum et al57 for a review). In surface dyslexia, the link between word forms and semantics is impaired so that reading occurs through phonology with characteristic errors for irregular words.58 The disorder might be considered a disconnection of regions specialised for word forms from regions specialised for visual semantics or a topological deficit of visual semantic specialisations.
The action aspect of depth vision (reaching behaviour) is dysfunctional in optic ataxia (impaired reaching with intact motor and visual function) and is thought to reflect an inability to integrate different coordinate frames within the parietal lobe.59 The failure of integration might reflect a hodological disconnection of the relevant reference frames or a topological deficit of an area specialised for their integration. A variant (crossed optic ataxia) in which reaching into contra-lateral hemispace is impaired (eg, reaching with the right hand for an object located on the left side) is thought to relate to the disconnection of the two hemispheres with a combination of splenial and parietal lesions.60
Oculomotor apraxia (a disorder of gaze in which subjects are unable to move gaze voluntarily from one object to another with preserved reflex gaze shifts) can be interpreted as a disconnection of voluntary saccadic systems in the frontal lobe from reflex saccadic systems in the parietal lobe (a network classified here as related to visual attention).
Denial of cortical blindness (Anton syndrome) might be considered a disconnection of visual cortex from body schema representations in the parietal lobe (classified here as related to visual attention). An inverse Anton syndrome (denial of seeing) has been interpreted as a disconnection of parietal attentional mechanisms and visual cortex.61 Denial of seeing specific objects reported in hypnosis (termed negative hallucination or scotomisation4) is likely to relate to the same mechanism.
In reduplicative disorders, a familiar person, place or object is perceived as replaced by a duplicate person, place or object62 (termed Capgras syndrome when involving a person). Reduplicative phenomena are thought to relate to the disconnection of visual and affective or memory regions.63 Similar disconnection accounts are given for derealisation (the feeling that experiences, typically visual, seem strange or unreal), visual hypoemotionality (where visual experience lacks emotional tone)64 and visual amnesia (a deficit of registering visual experiences in short-term memory).65
Visual alloaesthesia involves the transposition, rotation, tilting or inversion of the visual field (also referred to as reversal of vision metamorphopsia66) and has variants where the visual environment (but not the visual field) is rotated.2 The mechanism behind these disturbances is unclear but may relate to a disconnection of vestibular and visual inputs, transcallosal disconnection of visual and parietal inputs or topological lesions affecting regions specialised for the integration of these modalities.2 The disorders are associated with occipito-parietal lesions that spare the optic radiations and with brainstem pathology.66 Deficits of visual axis perception (eg, judgement of true vertical or horizontal) are associated with lesions of the anterior parietal lobe and have been attributed to a disconnection of vestibular inputs and their cortical representation or topological deficits of vestibular representations in the parietal lobe.67
The inability to mimic meaningless or meaningful actions or postures (visual imitative apraxia) is likely to reflect disconnection of visual and motor representations.68 Utilisation behaviour (automatic manual exploration and use of objects placed within the field of view and reach) has been interpreted as a deficit of frontal inhibitory function on parietal-led exploratory behaviour69 and might be considered a form of diaschisis. Automatic exploration of objects by placing them in the mouth forms part of the Klüver-Bucy syndrome (also including visual agnosia and hypersexuality) following anterior temporal lobe lesions.
Hyperconnection
Of the four classes of dysfunction, hodological hyperconnection is the most speculative; however, there is indirect evidence that connectivity increases are linked to perceptual symptoms in the auditory domain.70 Increased connectivity might lead to perceptual symptoms through, for example, enhanced feedforward or feedback processing or resonant activation through ephaptic (non-synaptic) transmission. The prototypical disorder in this class is synaesthesia. In the clinical domain, synaesthesia has traditionally referred to a percept in one sense modality evoking a parallel hallucinatory percept in a different sense modality—for example, percepts of taste evoking visual percepts of colour (coloured taste synaesthesia). More recently, work in the cognitive sciences has lifted the requirement of synaesthetic experience to cross modalities through the study of, for example, coloured grapheme synaesthesia (visual percepts of text evoking visual percepts of colours). The induced synaesthetic experience may be perceived in the external world (projector synaesthesia) or inside the mind's eye (associator synaesthesia).71 Other parallel sensory experiences recognised clinically include reflex hallucinations (a sensory stimulus in one modality evokes a hallucination in another, eg, seeing a butterfly whenever the thumb is touched) and functional hallucinations (hallucinations are triggered in the same modality as the inducing stimulus, eg, hearing voices in parallel with the sound of running water). Both clinical symptoms would be classed as synaesthetic by contemporary cognitive science. We would argue that the content of the parallel experience in such disorders is defined by the cortical specialisation of the area with increased connectivity. Thus, for example, synaesthesiae, reflex or functional hallucinations that involve colour relate to the connectivity of colour-specialised cortex, while experiences involving spatial location (ie, number forms—a variety of synaesthesia in which numbers and other ordinal lists, eg, days of the week, are located at specific spatial locations72) relate to the connectivity of spatial representations in the parietal lobe.
Pareidolia refers to images of objects or faces seen in random visual patterns (eg, cloud formations or tree bark). The experience is an illusion in which the inducing visual stimulus is typically of simpler form than the illusory percept and can be considered the result of hyperconnection between visual areas.
The Frégoli syndrome, in which unfamiliar people are perceived as familiar (typically as a person in disguise with malevolent intent), can be conceived as hyperconnection within visual emotional or memory networks. In post-traumatic stress disorder flashbacks, patients relive stressful events, typically with multiple sensory modalities involved and a strong emotional component. The flashbacks might be considered the result of hodological hyperconnection between visual and emotional systems or topological hyperfunction of visual memory systems.
The Perky effect originally referred to an ambiguity between visual imagery and visual perception such that normal subjects were unable to detect the presence of a real visual stimulus when asked to form a visual image of it. More recently, the term has been used to describe the reduction in visual perceptual sensitivity during visual imagery.73 The effect is likely to relate to connectivity between visual memory networks and occipital cortex. Eidetic imagery (sometimes referred to as photographic memory)74 is likely to relate to increased connectivity between these regions.
Visual phobias and visual objects appearing beautiful and comforting (kalopsia) or ugly and menacing (kakopsia)75 might be considered the result of hodological hyperconnection between visual emotional networks and occipital cortex or topological hyperfunction of visual specialisations within the amygdala.
Dazzle (painful brightness of visual stimuli) is thought to be a visual variant of a thalamic pain syndrome76 and can be interpreted as hodological hyperconnection of the thalamus and visual cortex. The same mechanism may account for the intense photophobia of children with cortical visual impairment and the paradoxical worsening of their visual acuity (as measured by the evoked response) for lighter (higher luminance) compared with darker (lower luminance) visual stimuli.77
Limitations
The classificatory scheme has a number of limitations and is perhaps better viewed as a structured review of symptoms from which to generate and test further experimental hypotheses than a definitive pathophysiological taxonomy or clinical aide memoire. First, it does not easily accommodate disorders involving combinations of networks or combinations of topological and hodological dysfunction (the latter marked with an asterisk in the table 1). Typical lesions have both hodological and topological effects so that pure disorders of one or other class may be largely theoretical. Second, the classification deconstructs traditional clinical syndromes by placing their component parts in different locations, losing associations between symptoms related to the anatomical proximity of the cortical regions underlying them. For example, in migraine aura, the same region of cortical spreading depression may result in a hyperfunctional symptom at its margins (teichopsia) and a deficit in its centre (a scotoma), but each is placed in a different location in the table 1. Similarly, the Balint syndrome (simultanagnosia, optic ataxia and oculomotor apraxia in association with biparietal lesions78) is spread across the table 1. Third, hyper- and hypofunction are convenient concepts to map on the statistical signal increases and decreases in imaging and tractography data. However, the neurophysiological meaning of such changes is unspecified. An increase in activity as measured by fMRI may have a number of neurophysiological causes (eg, a change in excitatory input or inhibitory processing, a shift from burst to tonic mode firing,36 neuromodulatory inputs, etc). Similarly DT-tractography dissects virtual connections between brain regions and different causes of microstructural white-matter change may lead to the same change in connectivity. By focussing on increases or decreases in imaging signal, the classification is likely to miss key pathophysiological distinctions. Finally, the classification does not easily accommodate interactions between predisposing factors and changes occurring at the time of a symptom. The Charles Bonnet Syndrome, for example, appears both as a hodological disconnection (because of the predisposing factor of loss of visual inputs from the eye) and as topological hyperfunction (because of the localised increases in fMRI signal during hallucinations).
Clinical implications
Despite its limitations, the classificatory scheme and its underlying assumptions are of relevance to current clinical practice in a number of areas. First, it suggests that, in the same way that seizure semiology is used in the context of epilepsy, the contents of hallucinations and illusions can be used as localising signs. By indicating increased activity in a specific functionally specialised cortical area, different types of visual hallucinations (eg, a face, colour or geometrical pattern) point to specific cortical locations, although not the location of the pathology causing the activity increase. Second, the classification highlights the fact that many symptoms held traditionally as distinct, appear closely related, if not indistinguishable, when considered from the perspective of their underlying mechanism. For example, synaesthesiae, functional hallucinations and reflex hallucinations are all likely to relate to the same underlying pathophysiology. Third, it predicts a number of theoretical disorders which are, as yet, undescribed, or at least not recognised as distinct clinical entities. For example, disorders classified here as hyperfunction of spatial coordinate frames within the parietal lobe (polyopia, visual perseveration, palinopsia, etc) do not have obvious deficit or disconnection counterparts. It is conceivable that their natural counterparts are, in fact, spatial neglect, simultanagnosia and oculomotor apraxia, classified here are as related to visual attention, as all are linked to the parietal lobe and dorsal stream. Finally, the classification highlights a number of areas which pose a challenge to visual neuroscience. For example, lilliputian hallucinations are not easily accommodated within current models of object perception, while polyopia and extracampine hallucinations are not easily accommodated within current models of spatial vision. In pointing to deficiencies in such models, visual perceptual disorders advance our understanding of normal visual function.
Conclusions
The last decade has seen advances in white-matter and functional imaging (including EEG imaging) that allow new classes of clinically relevant questions to be answered.79 We envisage that cutting-edge functional anatomical evidence derived from such techniques in single case and group studies, will be used to test and expand the classification presented here in the future.5 79 For now, we hope that our broad overview of visual perceptual disorders will provide a fresh perspective from which to challenge and refine the neuroscience of visual perception and visual perceptual dysfunction.
References
Footnotes
Linked articles 206516.
Competing interests None.
Provenance and peer review Commissioned; externally peer reviewed.
Linked Articles
- Editorial commentary