Article Text

Abstract
The fornix is a white matter bundle located in the mesial aspect of the cerebral hemispheres, which connects various nodes of a limbic circuitry and is believed to play a key role in cognition and episodic memory recall. As the most prevalent cause of dementia, Alzheimer’s disease (AD) dramatically impairs the quality of life of patients and imposes a significant societal burden on the healthcare system. As an established treatment for movement disorders, deep brain stimulation (DBS) is currently being investigated in preclinical and clinical studies for treatment of memory impairment in AD by modulating fornix activity. Optimal target and stimulation parameters to potentially rescue memory deficits have yet to be determined. The aim of this review is to consolidate the structural and functional aspects of the fornix in the context of neuromodulation for memory deficits. We first present an anatomical and functional overview of the fibres and structures interconnected by the fornix. Recent evidence from preclinical models suggests that the fornix is subdivided into two distinct functional axes: a septohippocampal pathway and a subiculothalamic pathway. Each pathway’s target and origin structures are presented, followed by a discussion of their oscillatory dynamics and functional connectivity. Overall, neuromodulation of each pathway of the fornix is discussed in the context of evidence-based forniceal DBS strategies. It is not yet known whether driving fornix activity can enhance cognition—optimal target and stimulation parameters to rescue memory deficits have yet to be determined.
- alzheimer's disease
- electrical stimulation
- neurosurgery
- limbic system
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
The fornix is a white matter bundle located in the mesial aspect of the cerebral hemispheres, which connects various nodes of a limbic circuitry and is believed to play a key role in cognition and episodic memory recall.1 2 While fornix lesions impair memory, it is not yet known whether driving fornix activity can enhance cognition. Stimulating a distributed fibre bundle such as the fornix could be a powerful and efficient method of targeting interconnected brain regions involved in cognitive processes.2 3 Importantly, brain-wide targets could be simultaneously modulated by stimulation of forniceal fibres projecting from and to widely distributed regions. A detailed knowledge of the neuronal structures interconnected by the fornix bundle would be crucial to delineate the antidromic and orthodromic effects of neuromodulation. A comprehensive knowledge of the forniceal anatomy could also serve to inform clinicians to cognitive subpathways and corresponding memory deficits that arise from neurodegenerative disease or lesions. Likewise, careful consideration of structures targeted by forniceal projections could also inform the electrophysiologist of optimal deep brain stimulation (DBS) parameters to try to rescue particular types of memory deficits encountered in Alzheimer’s disease (AD), traumatic brain injuries, epilepsy or strokes.
Neuromodulation of the fornix by electrical DBS has recently been investigated as a treatment for memory impairment in patients with AD.4 In a phase I study, one-third of patients showed improvement or slowing in the rate of cognitive decline. In a recent phase II study, it was suggested that the subgroup of patients with AD over the age of 65 tended to experience a slower deterioration in memory, and a phase III trial is currently ongoing to study the neuropsychological effects of fornix DBS in this patient subgroup.5 Despite multiple clinical trials underway, optimal target and stimulation parameters to rescue memory deficits have yet to be determined, and the therapeutic benefit is modest.2 5
Here, we review structural and functional aspects of the fornix in the context of neuromodulation for memory deficits. We suggest that the fornix should be subdivided into two distinct anatomo-functional axes: a forniceal septohippocampal (SHP) pathway and a forniceal subiculothalamic pathway. Each pathway’s target and origin structures will first be presented, followed by a discussion of their oscillatory dynamics and functional connectivity. In the context of evidence-based therapeutic forniceal DBS strategies, neuromodulation of each axis will be discussed with supporting preclinical and clinical evidence.
Gross anatomy of the fornix
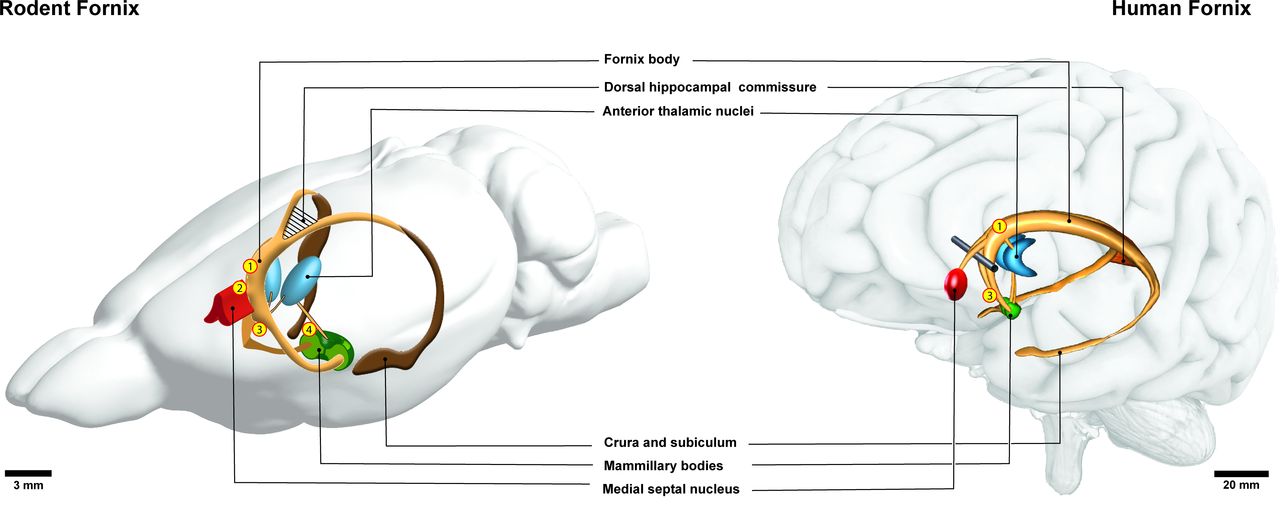
The fornix is a thin arched white matter bundle composed of myelinated association, projection and commissural fibres located in the mesial aspect of the cerebral hemispheres (figure 1). The human fornix contains approximately 1.2–2.7 million fibres in each hemisphere6 7 and fills a total volume of about 1000–1800 cubic millimetres.8 9 As a major hippocampal output structure, the fornix stretches longitudinally from the mesial temporal lobe to the diencephalon and basal forebrain. Medial to the floor of the temporal horn of the lateral ventricle, hippocampal fibres collect into a thin lamina known as the alveus. Fibres from the subiculum join the alveus as it courses posteromedially and bundles into the fimbria of the fornix. As the fimbria enlarge in cross-sectional area by collecting additional fibres, they become known as the crura of the fornix. The crura arch supero-anteriorly under the splenium of the corpus callosum and project contralaterally via the thin triangular forniceal commissure, also known as the psalterum or dorsal hippocampal commissure. The crura run paracentrally to form the forniceal body, which arches over the thalamus and under the septum pellucidum. Rostrally, the fornix body bifurcates into left and right columns that descend into the basal forebrain anterior to the interventricular foramina. The fornix columns divide at the anterior commissure—fibres travelling anteriorly form the pre-commissural fornix, while those curving posteriorly make up the post-commissural fornix. This division in structure reflects the two major forniceal functional pathways. Pre-commissural fibres house the septohippocampal pathway, also projecting to the forebrain. Post-commissural tracts originate from the subiculum and project to the thalamus, forming the direct subiculothalamic pathway, and the indirect subiculothalamic pathway which relays via the mammillary bodies (figure 2). Important cross-species differences exist between primate and rodent fornices, in part due to the distinct spatial configuration of their respective hippocampi. The rodent hippocampus is transposed more rostrodorsally, with its dorsal and ventral components hinged by a 90-degree flexure10 and envelops the hippocampal formation as a sheet of fimbria and alveus fibres.11 Compared with primates, rodents have a more developed commissural system—their dorsal commissure spans almost the entire longitudinal axis of the fornix. In addition, a thin transverse lamina known as the ventral hippocampal commissure (VHC) is found in non-human primates and rodents just ventral to the columns at the level of the subfornical organ.12 13 The VHC carries decussating dentate gyrus fibres14 and is probably absent in humans, though histopathological evidence exists of decussating fibres in this region15 16 which later studies were unable to reproduce.17 The dorsal fornix is the murine homologue to the human fornix body and its fibres arise from the temporal hippocampal pole and course along the undersurface of the corpus callosum, medial to the septal hippocampus.18 The thin dorsal fornix eventually disappears at the anterior commissure, where fibres of the rodent fimbria–fornix divide to reach their terminal nuclei.19

Gross anatomy of the rodent (left) and human (right) fornix. Locations where deep brain stimulation has been performed in rodents and humans are indicated by numerals—1: stimulation of the post-commissural dorsal fornix; 2: stimulation of the pre-commissural fornix, 3: stimulation of the post-commissural ventral fornix; 4: stimulation of the mammillothalamic tract. The results associated with these sites of stimulation are detailed in tables 2–5.

Simplified neurochemical anatomy of the fornix highlighting the presence of a septohippocampal pathway and a subiculothalamic pathway. The fornix is composed of neural populations comprising GABAergic, glutamatergic and cholinergic fibres. Septohippocampal projections encompass slow-firing (0.5–5 Hz) cholinergic, fast-firing and burst-firing (10–18 Hz) GABAergic and glutamatergic neurons. The subiculothalamic pathway comprises chiefly glutamatergic neurons projecting to the mammillary bodies and the anterior thalamic nuclei. AC, anterior commissure; DG, dentate gyrus; ATN, anterior thalamic nuclei; MB, mammillary bodies; MS, medial septum.
Forniceal pathways and memory impairment
Lesions of the fornix
In early literature, surgical lesioning of the anterior fornix for treatment of epilepsy was only rarely associated with subsequent memory deficits.20 More recently, cognitive deficits in episodic memory are increasingly being reported in patients with injuries to the fornix (table 1).21–23 Bilateral lesions of the fornix anterior columns are associated with anterograde and retrograde amnesia.24 25 The fornix carries distinct functions depending on laterality; the left fornix primarily carries verbal memory information, while the right carries visuospatial memory information.26–28 In addition, the medial fornix carries fibres from the caudal hippocampus, which integrates object recognition within a spatial context such as in scene learning. The lateral aspect of the fornix is thought to carry projections from the more rostral hippocampus, which processes interoceptive signals for emotional and motivational learning and memory.29 30 After forniceal lesions, a relative sparing of recognition is seen when compared with the loss of the recall of episodic information.23 27 Moreover, as nodes of the fornix, the mammillary bodies are classically considered to provide a relay for indirect hippocampal inputs to the anterior thalamic nuclei. In such models, the mammillary bodies do not have an independent role in memory. However, the medial aspect of the mammillary bodies’ contribution to memory is more dependent on afferents from the ventral tegmental nucleus of Gudden than from the hippocampal formation, while the lateral mammillary bodies, connected with the dorsal tegmental nucleus of Gudden, participate in generating head-direction signals.31
Studies examining the effects of fornix lesions in rodents, non-human primates and humans, along with associated behavioural deficits
Early Alzheimer’s pathology and the fornix
The neuropathological hallmarks of AD, such as extracellular beta-amyloid and intracellular tau, can provide insight into the neuroanatomical progression of the disease and shed light into potential neuroanatomical hotspots for neuromodulation-based therapy.32 33 The appearance of hyperphosphorylated cytoskeletal tau within the brain is found even before cognitive deficits are clinically apparent (ie, Braak and Braak AD stages 0 and 1), classically manifesting first in the entorhinal and transentorhinal regions.34 35 However, recent pathoanatomical studies in brains from cognitively intact individuals have found evidence of immunoreactive neuronal tau cytoskeletal pathology in subcortical nuclei interconnected by the fornix.34 35 Areas with presence of tau deposits in Braak stages 0 or 1 included the perifornical and lateral region of the hypothalamus, the dorsomedial, ventromedial, tuberomammillary and supramammillary nuclei, and subnuclei of the amygdala and thalamus known to be synaptically connected to the hippocampus via the forniceal and entorhinal pathways.34 Since PET imaging studies point to the distribution of tau signal as a strong predictor of future local neurodegeneration and atrophy, neuromodulation of the fornix or its subnuclei may represent a logical strategy.33 Indeed, studies in animal and in vitro models of AD have shown that synaptic activation via chronic DBS reduced pathological tau and provided synaptic neuroprotection.36 37
In addition to patterns of tau accumulation, another biomarker of AD correlating with disease severity includes alterations in cortical EEG dynamics.38 As a cortical dementia, hallmarks of underlying neuropathological changes in AD include decreases in lower cortical frequency bands (alpha and beta), and an increase in theta and delta rhythms.38 39 Reduced coherence of cortical alpha and beta bands is also seen in AD, suggesting losses in cortical synaptic function.38–40 Since the fornix is a deep structure connecting chiefly subcortical nuclei, at the time of this writing no DBS studies have studied cortical EEG changes in response to DBS, though it merits future study as a potentially useful outcome measure to monitor response to neuromodulation treatment.
Forniceal connectivity and memory impairment
Functional MRI studies are beginning to suggest that degeneration of the fornix bundle itself may precede hippocampal dysfunction and predict cognitive impairment better than structural measures such as hippocampal atrophy.41 Connectivity data from diffusion tensor imaging studies suggest that forniceal measures correlate with episodic memory performance in various neuropathological conditions, as well as during brain development and ageing. On imaging, macrostructural and microstructural alterations of the fornix have been found to be robust predictors of episodic memory performance, independent of age and associated structural pathology.1 42 Specifically, in AD, atrophy of the fornix on structural MRI and reductions in fractional anisotropy have often been reported.43–45 Fornix atrophy may predict the onset of AD,41 even prior to clinical manifestations. Moreover, fornix fractional anisotropy reduction is correlated with cognitive decline in AD.46 While fornix lesions or degeneration are associated with memory impairment, it is not yet known whether driving fornix activity can enhance these functions.
Septohippocampal pathway
Anatomy and spontaneous activity
The projections from the medial septum (MS) to the hippocampus form the septohippocampal forniceal pathway and are proposed to have important roles in cognition by modulating the activity of episodic memory circuits.47–49 As the terminal structure of this forniceal pathway, the hippocampus is necessary for episodic memory, and is involved in the storage and recall of autobiographical events.50 51 Sensory cue inputs from the entorhinal cortical grid cells and memory-related internal brain activities govern the firing of hippocampal neurons.52–55 Traditionally, the hippocampus has been thought to exhibit two dominant and behaviour-dependent local field potential (LFP) patterns: theta rhythm and large-amplitude irregular activity with sharp waves.56 Theta rhythm is a large-amplitude (1–2 mV) 4–10 Hz sinusoidal oscillation in the rat,57 58 with two defined subtypes: type 1 (7–10 Hz) is associated with voluntary movement and exploratory behaviour, whereas type 2 (4–6 Hz) is present during immobility, rapid eye movement (REM) sleep or urethane anaesthesia.59
Theta oscillations are understood to be critical in hippocampal mnemonic and learning functions.58 60 –66 The MS projections through the fornix are involved in hippocampal theta modulation.47 ,62,67,68 Rhythmically discharging cells of the MS in the diagonal band vertical nucleus fire synchronously with theta and may be involved in its pacing.58,67,69–76 In freely moving rats, MS neuron activity can be negatively (during sharp wave ripples) or positively (during theta waves) correlated with the activity of hippocampal neurons.75 The MS decreases the spiking of hippocampal pyramidal neurons and reduces their ability to fire in trains.49 ,77 Septohippocampal oscillatory regulation of neuronal activity also precisely synchronises postsynaptic potentials arriving at hippocampal pyramidal cells.78
Conversely, disrupting or lesioning the MS eliminates the hippocampal theta rhythm. During MS inactivation by muscimol79 or lidocaine,77 grid cells recorded in the medial entorhinal cortex lose their spatial periodicity. Also, loss of theta via MS lesioning significantly alters performance on spatial49,80,81 as well as non-spatial tasks.80,82 However, after extinguishing hippocampal theta rhythm via pharmacological inactivation of the MS,77,80 location-specific firing of hippocampal place cells are maintained. Sensory cues seemingly guide hippocampal neural firing in rats, whereas MS inputs prevail over shorter timescales,83 and both support the formation of hippocampal spatial firing fields.
In rats, reciprocal hippocamposeptal projections exist to the cholinergic nuclei of the MS via forniceal GABAergic neurons.84,85 The CA1 to MS fibres are sparse and project unilaterally, whereas those originating from CA2–3 project extensively and bilaterally.86 Topographically, the dorsal CA3 innervates the dorsal and medial parts of the MS; conversely, axons of the ventral CA3 reaches the lateral and ventral parts of the MS.86 The hippocamposeptal tract has been implicated in inhibiting87,88 and modulating88 theta generators in the septal region.85 For instance, in vitro rodent hippocampal preparations demonstrated the role of hippocamposeptal modulation in phasing the spiking of MS GABAergic neurons, while inhibiting acetylcholinergic and glutamatergic neurons in the same region.89
Electrical stimulation of the septohippocampal pathway
Diverse stimulation parameters of the SHP have been explored. High-frequency (100 Hz) chronic or acute stimulation of the rodent SHP were found to induce hippocampal long-term potentiation and neurogenesis.90 Furthermore, upregulation of genes involved in synaptic function, cell survival and neurogenesis was observed in molecular expression studies after such SHP stimulations.91
Because theta oscillations are critically involved in memory, SHP stimulation at theta frequencies has been thoroughly investigated in rodents (figure 1; table 2). Specifically, electrical stimulation of the MS in the 3–12 Hz range serves to experimentally mirror physiologic theta-like hippocampal LFP frequencies.92–95 Indeed, 5–7 Hz DBS exhibits electrophysiological characteristics similar to those of spontaneously occurring theta field activity,96 while higher stimulation frequencies produce hippocampal desynchronisation.97–100 Although CA1 pyramidal cells respond maximally to 6–8 Hz MS stimulation, electrical stimulation of medial septal nuclei does not produce typical physiological hippocampal theta-related activity.101 Nevertheless, restoring theta-like hippocampal activity by stimulation of the SHP was shown to rescue memory deficits in rats after MS inactivation.95 More precisely, irregular SHP stimulation resulted in little rhythmicity, while a fixed stimulatory frequency of 7.7 Hz triggered by supramammillary nucleus theta rhythmicity restored theta-like rhythmicity with abnormal waveforms. While both stimulation paradigms improved memory deficits, the latter was the most efficient. Thus, despite incomplete physiological reinstatement, promotion of synchronous low-frequency phasic firing rescues learning processes in rodent models. DBS of the SHP has also been shown to restore cognitive deficits associated with pathological brain states. For instance, in a rat model of traumatic brain injury, theta stimulation of the MS restored hippocampal theta oscillations and yielded improvements in object exploration when performed chronically,102 as well as improved spatial working memory when administered acutely before training.103
Stimulation of the post-commissural dorsal fornix in rodents and patients with epilepsy, along with main findings
Moreover, temporal co-ordination of theta and gamma rhythms is important for sequential memory retrieval,104,105 sequential representations during learning,106,107 and facilitation of synaptic plasticity.108,109 After pharmacological inactivation of the MS, Shirvalkar and colleagues reported that acute theta burst stimulation (TBS) of the SHP within the fimbria–fornix region increased hippocampal theta–gamma coupling (TGC) in amnestic animals and rescued memory performance in the Morris Water Maze.110 Notably, single-trial spatial memory performance in rats was predicted by the power comodulation of theta (4–10 Hz) and low gamma (30–50 Hz) rhythms in the hippocampus during the retrieval phase. However, TGC was weak when memory failed and was unavailable during spatial exploration. Thus, TGC may be necessary for memory encoding and retrieval.
TBS is a distinct pattern of stimulation that has been investigated for its physiological relevance. Non-selective septal TBS resets hippocampal theta cell bursting during active behaviour while increasing theta synchronisation.96,111–115 This reset persists for 600–900 ms and enables dentate granule cell depolarisation at the time of sensory input arrival from the entorhinal cortex. This synchrony facilitates long-term potentiation, enabling synaptic plasticity,116,117 and ultimately enhancing the encoding of incoming information.118–121 TBS also entrains the spiking of hippocampal place cells,115 potentiates population spikes at CA1122 and temporally regulates the place field spatial properties during active exploration.115
MS neuronal subpopulations
Septohippocampal projections encompass immunohistochemically and electrophysiologically distinct slow-firing (0.5–5 Hz) cholinergic, fast-firing and burst-firing (10–18 Hz) GABAergic and glutamatergic neurons with heterogeneous firing patterns123–138 (figure 2). Co-synthesis of glutamate in cholinergic and GABAergic neurons has also been reported.131,133 Connections include sparse GABAergic inputs from lateral to medial septal cholinergic neurons, reciprocal connections between medial septal cholinergic and GABAergic neurons, and also glutamatergic neurons within the MS synapsing onto neighbouring cholinergic and GABAergic neurons.115,139–141 In primates, the MS projects to the hippocampus in a topographically oriented fashion: medial portions project via medial fornix fibres, and the lateral MSN project via the lateral fornix.29,142 To understand the physiology of the rodent SHP, the projection patterns for each MS neuronal subtype will be reviewed.
MS GABAergic neurons
Connectivity
Medial septal GABAergic fibres terminate on vasoactive intestinal polypeptide–immunoreactive interneurons in strata pyramidale and lacunosum-moleculare of the CA1143 and on calretinin-immunoreactive and neuropeptide Y–immunoreactive GABAergic interneurons in the stratum radiatum of the CA1 and stratum lucidum of CA3.127,144–146 Inhibitory inputs terminate on neurons containing cholecystokinin, somatostatin and parvalbumin in the stratum oriens.127,144,146,147
Electrophysiology
GABAergic septohippocampal projection cells are crucial for hippocampal theta generation.59 ,67,74,75,148–151 For instance, the frequency of theta in urethane-anaesthetised rats depends on the activity of medial septal GABAergic cells.152 GABAergic neurons may pace the hippocampus in vivo via tonic, cluster or burst-firing patterns of discharge mediated by low-threshold spikes and prominent hyperpolarisation-activated cation (Ih) currents.135,136,153 Rhythmic bursting activity, observed at theta frequencies during wakefulness and REM sleep, is more pronounced in GABAergic neurons that contain parvalbumin. These neurons display higher discharge rate and longer burst duration,136,154–156 which may result from the calcium-buffering properties of parvalbumin.138 Local GABAergic MS neurons typically do not contain parvalbumin.157 Burst-firing neurons tonically fire during slow-wave sleep, and their discharge rates remain high across the sleep/wake cycle. Rhythmic bursting activity at theta frequencies is tightly coupled to hippocampal theta waves: glutamic acid decarboxylase (GAD)–positive neurons are distributed in almost equivalent proportions between T type (burst-firing at trough of hippocampus theta) and P type (peak of hippocampus theta) whereas the parvalbumin/GAD+ are largely T type.136,154,155,158 Furthermore, the hyperpolarisation-activated and cyclic nucleotide-gated non-selective cation channel (HCN) of some MS GABAergic neurons has been suggested to play a pacemaker role for theta oscillations154,156 by disinhibiting the hippocampal pyramidal cells via rhythmic inputs to hippocampal GABAergic interneurons.127,159 Activity changes within parvalbumin and/or HCN neurons precede changes in hippocampal interneurons and theta rhythm.158
Behaviour
An important component of hippocampal network dynamics and plasticity during learning160 is the modulation of CA1 interneuron activity by the septohippocampal GABAergic pathway during salient sensation and locomotion. Furthermore, Cav3.1 T-type Ca2+ channels are highly expressed in the septohippocampal GABAergic projection neurons161 and are critically involved in controlling object exploration through modulating hippocampal type 2 theta rhythm.162 Specifically, optogenetic activation of this pathway in mice selectively enhances novel object exploration and type 2 theta rhythm, whereas inhibition of the same pathway decreases both exploration and the rhythm.162
MS cholinergic neurons
Connectivity
Medial septal cholinergic terminals project to all regions of the hippocampus,163,164 especially the stratum oriens of the CA1 and CA3 subfields.165–167 These terminals synapse with pyramidal cell dendrites168 and cell bodies and dendrites of GABA-containing and somatostatin-containing interneurons147,166,169,170 and dentate granule cells.171 As the majority of axon terminals are diffusely organised172 and do not associate with distinct postsynaptic sites,165,166,168,173 cholinergic transmission in the hippocampus is likely primarily mediated by volume transmission. The cholinergic projections to the hippocampus may therefore tonically maintain an extracellular ambient level of acetylcholine,174 leading to long-lasting effects.173,175
Acetylcholine release within hippocampal circuits results in the activation of both metabotropic muscarinic (mAChRs) and ionotropic nicotinic (nAChRs) acetylcholine (ACh) receptors. Nicotinic receptors are expressed in dentate granule cells, pyramidal cells and interneurons both presynaptically and postsynaptically,176 on interneuron axons terminating on excitatory and inhibitory neurons177–181 and at inhibitory synapses contacting pyramidal neurons.182 Muscarinic receptors are expressed in soma and dendrites of pyramidal neurons and granule cells, with a small fraction expressed on axons and terminals.183 These receptors are also found in interneurons184,185 and in fibres surrounding pyramidal cells. The highest density of expression is found presynaptically in GABAergic terminals projecting onto the perisomatic region of pyramidal cells,186–189 and postsynaptically in dendrites and cell bodies of interneurons in the stratum oriens and alveus of CA1,190 or in glutamatergic terminals.187
Electrophysiology
As a consequence of their presynaptic and postsynaptic location, muscarinic receptors can have diverse impacts on hippocampal neuronal activity, influencing the net effect of ACh. Choline acetyltransferase (ChAT)–positive neurons have a long duration spike136 and fire at a lower frequency136,191 during the inactive (3.4±0.3 Hz) compared with active behavioural state (4.7±0.3 Hz).115 ACh has an excitatory effect on GABAergic and glutamatergic neurons within the MS.141,192,193 Cation flux through nAChRs mediates fast excitatory synaptic responses.179,181,182,194–199 Fast membrane depolarisation triggers activation of voltage-gated Ca2+ channels, second messenger systems involving cAMP200,201 and release from intracellular stores.202,203 Moreover, nAChRs may modulate pre-existing oscillatory states204,205 by enhancing a slow calcium-dependent potassium conductance that reduces the firing of stratum oriens interneurons.206 In contrast to the fast response produced by activation of nAChRs, mAChR-mediated transmission is slow, owing to their dependence on G-protein-coupled signalling mechanisms.207 Moreover, M1/M3 mAChR activation sharpens interneurons’ firing precision to theta frequency input, leading interneurons to amplify theta oscillations.208
ACh can suppress or enhance presynaptic neurotransmitter release in the hippocampus.178,179,182,188,189,209–215 ACh can facilitate and induce hippocampal long-term potentiation (LTP) or depression (LTD).128,199,216–225 The precise mechanism and direction of modulation may depend on ACh concentration, the timing of its release, exposure time, and the temporal sequence of nAChR and mAChR activation in relation to ongoing neuronal activity.199,226–228
Cholinergic MS neurons can be selectively activated by optogenetics.115 The evoked hippocampal response involves direct activation of ChAT projections together with indirect activation of non-ChAT septal neurons. Hippocampal neurons respond with an initial inhibition followed by rebound potentiation, inhibition and biphasic response, including potentiation and subsequent inhibition. Optogenetic septal ChAT stimulation exerts frequency-dependent and behaviour-dependent effect on hippocampal formation. The spiking of hippocampal neurons is significantly increased by 50 Hz but not 10 Hz septal stimulation and spiking increase is higher for inactive behavioural state than for active behavioural state.
Although cholinergic neurons are not pacemakers for hippocampal theta oscillations, they are involved in the generation and modulation of some of their attributes.59 ,62,67,74,148–150,229,230 However, the long post-hyperpolarisation period, small Ih and slow firing rates characteristic of MS cholinergic neurons limit their capacity to pace theta-related rhythmically bursting activity.126,128,135,136,231 Moreover, selective lesioning of MS cholinergic neurons reduces the number of rhythmically bursting neurons in the MS232 but does not eradicate hippocampal theta.151 Cholinergic MS neurons have been shown to display very slow, theta-unrelated firing in vivo, suggesting that these neurons might not act as pacemakers but rather generate theta activity via the tonic excitation of MS GABAergic and glutamatergic neurons136,141,192 and hippocampal interneurons and principal cells.166 Although in vivo coupling between phasic ACh release and theta oscillations has been shown, theta initiation was found not to require Ach.191 Cholinergic neurons firing was also seen to follow theta oscillations, making them pro-arousal and not pacemaker neurons for hippocampal theta oscillations.191
Cholinergic neurons also modulate the amplitude of theta oscillations. Microdialysis of ACh release152 and selective lesions of septohippocampal cholinergic neurons233 showed that cholinergic neurons selectively modulate the amplitude of theta oscillations, and not its frequency.136,234 Optogenetic activation of MS cholinergic neurons affects hippocampal oscillations in the theta range in a behaviour-dependent and frequency-dependent manner. Theta and gamma both significantly increase for inactive compared with active rodents115,234 and for low-frequency stimulation (8–10 Hz).115 The overall tone of ACh is lower under urethane anaesthesia than in the awake animal235,236 and the faster theta seen during movement is largely non-cholinergically mediated, whereas the cholinergic input provides a background of sensory input–related drive to the hippocampus during movement.100 Another robust effect of optogenetic activation of MS cholinergic neurons in both anaesthetised and behaving mice is the suppression of slow oscillations in the supratheta frequency band.234
Cholinergic neurons have been found to play a role in hippocampal theta and synaptic plasticity. For instance, the local field synchronisation expressed a preference for a low-frequency stimulation protocol, whereas the hippocampal neuronal response showed significant increase after 50 Hz but not after 10 Hz optogenetic septal stimulation.115 While slow-spiking septal cholinergic neurons are linked to the amplitude of theta rhythm233 by tonically depolarising pyramidal cells and basket interneurons,237 the fast-spiking septal GABAergic cells are linked to the frequency of theta rhythm by periodically hyperpolarising hippocampal basket cells and rhythmically disinhibiting the pyramidal cells.159
Behaviour
Optogenetic cholinergic stimulation of the MS does not exert a consistent effect on locomotion velocity and motor behaviour.114,115,234 In fact, inhibition from different classes of interneurons create gamma oscillations within each theta cycle, and the modulated efficacy of excitatory inputs at different theta phases can selectively influence the timing of pyramidal cell firing.238 Therefore, promotion of co-ordinated firing and rhythmic activity by ACh release may provide an increase in the baseline excitability of neurons. This results in enhanced neural responses to glutamate and promotes neural interactions facilitating memory formation. Within this system, synaptic input that arrives during the positive phase of theta induces LTP while input that occurs during the negative phase induces LTD or depotentiation.239–243 In addition, cholinergic receptor activation enhances LTP induction during exploration218 and theta entrained hippocampal place cell activity.222 Therefore, high cholinergic tone during phases of exploration, in synergy with optimally timed theta, enhances plasticity.
MS glutamatergic neurons
Connectivity
Some glutamatergic neurons within the MS provide functional excitatory input to local cholinergic and GABAergic neurons.132,140,141,244 Others send direct projections to a restricted number of pyramidal cells and interneurons in the hippocampus.137,245
Electrophysiology
Medial septal glutamatergic neurons expressing type 2 vesicular glutamate transporters (VGluT2) are likely involved in hippocampal theta generation.132,135,140 They display a heterogeneous firing pattern, including fast, slow, burs, and cluster-firing (8–14 Hz, half of glutamatergic neurons) properties in slice.135,137 Glutamatergic neurons also have intrinsic firing properties that may play an important role in pacing the hippocampus in vivo: they can discharge in recurrent clusters of action potentials, interspersed with intrinsically generated subthreshold membrane potential oscillations.135
Behaviour
Before the onset of locomotion, the hippocampus undergoes a transition into an activity-state specialised for the processing of spatially related input. This brain-state transition is associated with increased firing rates of CA1 pyramidal neurons and the occurrence of theta oscillations, which both correlate with locomotion velocity.56 ,62,246–250
Fuhrman et al showed that the SHP mediated by glutamatergic (VGluT2+) neurons is activated before locomotion onset and controls the initiation and velocity of locomotion as well as the entrainment of theta oscillations.251 Septohippocampal projections onto alveus/oriens interneurons regulate inhibition of Schaffer collateral and perforant path input to CA1 pyramidal neurons in a locomotion-dependent manner. Accordingly, with faster locomotion, the increased activity of medial septal VGluT2 neurons is translated into increased axo-somatic depolarisation and higher firing rates of CA1 pyramidal neurons. Conversely, rhythmic activation of septal glutamatergic neurons can powerfully drive hippocampal theta oscillations through local septal interactions with GABAergic and cholinergic neurons251,252 rather than through direct glutamatergic projections to the hippocampus.252 Indeed, Robinson et al found that glutamatergic neurons provide prominent excitatory inputs to a majority of local GABAergic and a minority of septal cholinergic neurons.252
In contrast, activation of MS glutamatergic fibre terminals in hippocampal slices elicited weak postsynaptic responses in hippocampal neurons. Thus, MS VGluT2 neurons may couple neuronal firing rates and theta oscillations to movement velocity. Moreover, Fuhrman et al showed the in vivo role of MS VGluT2 neurons in the pre-motor control of locomotion.251 Firing of MS VGluT2 resulted in reliable initiation of locomotion, and also the firing rate and number of activated VGluT2 neurons directly correlated with the speed and the duration of locomotion. The pre-motor activity of MS VGluT2 neurons predicted the speed and delay to onset of the upcoming movement. In parallel, the pre-motor activation of MS VGluT2 neurons led to reliable entrainment of hippocampal theta oscillations before locomotion onset. Furthermore, Furhman et al raised the possibility that a small fraction of VGluT2-positive septohippocampal axons may directly excite the dendritic compartment of pyramidal neurons.251
Together, these findings strongly suggest an important role of the MS VGluT2 circuitry in the concerted initiation of locomotion and theta oscillations and in the active regulation of locomotion speed.
Septohippocampal pathway summary
Overall, MS burst-firing GABAergic neurons are important in generating, maintaining and pacing hippocampal theta activity by modulating GABAergic hippocampal interneurons and, indirectly, pyramidal cells.127,151,158,159,253 In turn, slow-firing cholinergic cells modulate theta amplitude.232,233 Septal glutamatergic neurons provide dense connections within the septum and comparatively sparse projections across the hippocampus: their rhythmic activation can powerfully drive hippocampal rhythms through local septal interactions rather than through direct projections to the hippocampus.252
Furthermore, spontaneous activity of MS neurons can be influenced by different inputs from the locus coeruleus, raphe nuclei and hypothalamus.254,255 Therefore, in addition to acting as one of several extrinsic rhythm generators that work in concert to amplify and regulate intrinsic theta generators within the hippocampus, the MS may relay and pace theta rhythm by integrating inputs from neighbouring brain regions.256 Taken together, a neurophysiologically-inspired septal DBS protocol should combine at minimum a low-frequency cholinergic-like and high-frequency GABAergic-like stimuli.
Subiculothalamic pathway
Subicular projections to the anterior thalamic nuclei (ATN) and the mammillary bodies (MB) connect distant nodes of the circuit of Papez and propagate the theta rhythm generated by the MS. To understand the role of these projections, anatomy, physiology and functions of these nodes will be explored.
Subiculum
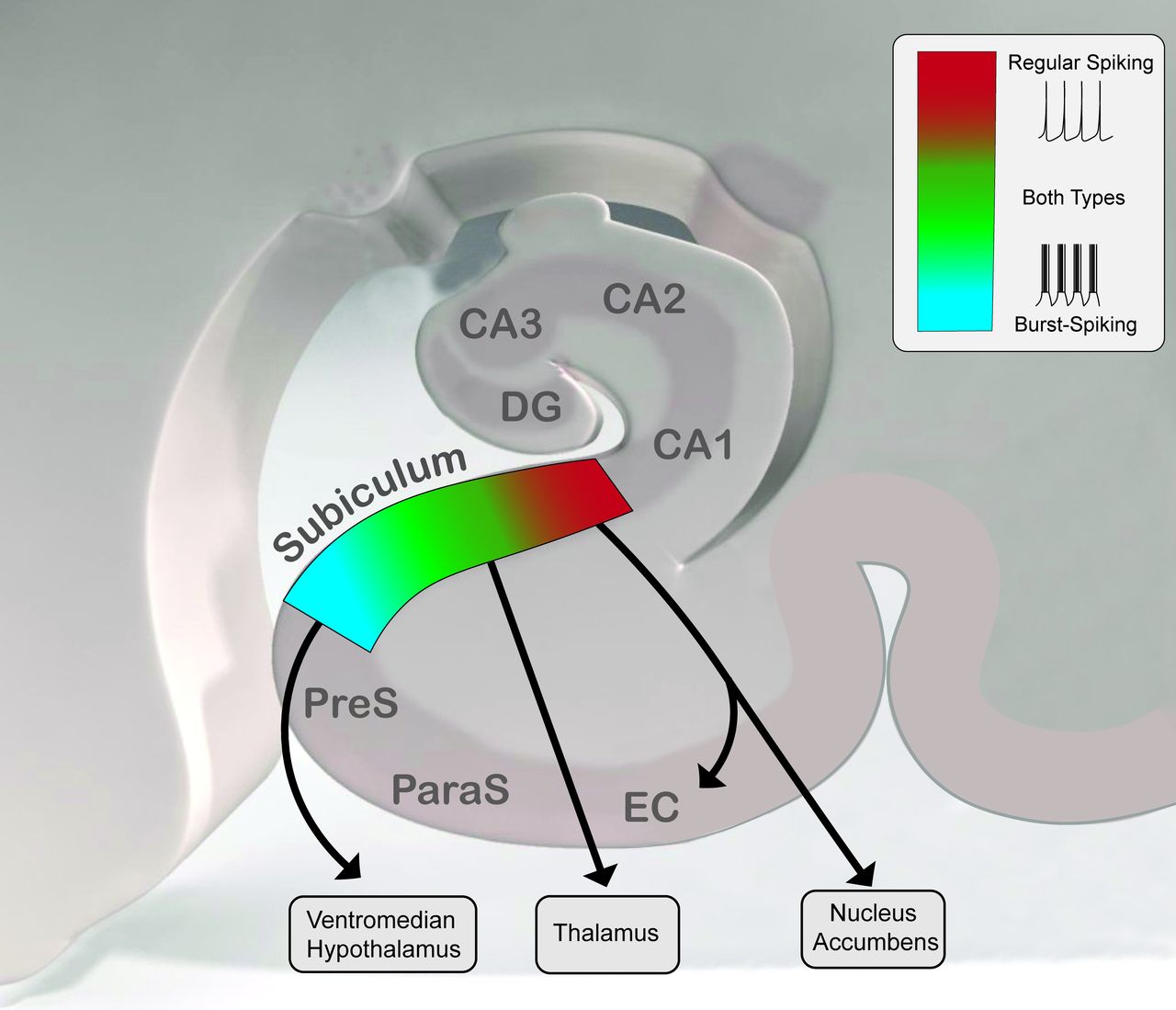
The subiculum constitutes the major output structure of the hippocampal formation.257 As the final relay in a polysynaptic loop between the entorhinal cortex (EC) and the hippocampus, it integrates and distributes processed spatial and mnemonic information to cortical and subcortical brain regions.258 Pyramidal neurons, which form the chief output of the subiculum, are divided into two main groups based on their firing mode: regular-spiking and bursting neurons (figure 3).259–263 Regular-spiking neurons fire a single action potential with 60–160 ms interspike intervals, whereas bursting neurons emit 2–5 action potentials at high frequency (2–5 ms interspike intervals) with decreasing successive spike amplitudes260,262,263 followed by a 20–30 ms refractory period with subsequent return to spiking. Bursting neurons are better suited to discriminate the content of high-frequency input, such as that occurring during gamma oscillations, than regular-spiking neurons.262 Interneurons are also found in the subiculum, as fast spiking units with an inter-spike interval of 7–10 ms and small spike width (<0.2 ms). There are more bursting than non-bursting neurons in the subiculum and both groups are distributed in an organised fashion along the proximal–distal axis, with more regular-spiking neurons close to CA1, and more bursting neurons close to the presubiculum.260,262 Moreover, there are distinct output targets in different portions of subiculum.257,258,264–268 The subiculum can be divided in four regions following the dorso-ventral and the proximo-distal axes, each of which serves as the origin of different parallel efferent projections with very few collateralisation.265,269–271 Bursting and regular spiking cells mainly target mainly respectively the presubiculum and the EC.272 Neurons projecting to the nucleus accumbens are located in the proximal subiculum and consist mostly of regular-spiking neurons (~80%) whereas neurons projecting to the ventromedial hypothalamus are located in the distal subiculum consisting mostly of bursting neurons (~80%), and neurons projecting to thalamus are located in the middle portion of subiculum with a bursting probability of 50%.273 Differences in the distribution and projection of regular-spiking and bursting neurons suggest that different types of information are conveyed from the subiculum to its various targets. Furthermore, the subiculum is capable of intrinsically generating two major memory relevant network rhythms: SWR274 and gamma oscillations.275 Gamma activity arises after tetanic stimulation of the subiculum or the hippocampal CA1 region.276,277 SWR might mediate memory consolidation and gamma oscillations the encoding of new information. A relatively small proportion of subicular recordings are phase-locked to theta. Nevertheless, similar to the hippocampus, subicular EEG is characterised by theta oscillations dominating exploratory behaviours, while SWR occur mainly during alert, still and quiet behaviours.
{kind=link}
{kind=link}
{kind=link}
Topographical organisation and spatial projection of regular-spiking and burst-spiking neural subpopulations within the subiculum. Pyramidal neurons are divided into two groups based on their electrophysiological properties: regular-spiking and bursting neurons. Regular-spiking neurons fire with 60–160 ms interspike intervals, whereas bursting neurons fire at high frequency with decreasing successive spike amplitudes. EC, entorhinal cortex; PreS, presubiculum; ParaS, parasubiculum; DG, dentate gyrus.
Several studies have investigated subicular functional connectivity and plasticity.278 In 2000, Gigg and colleagues showed that stimulation of CA1 produced excitation–inhibition sequences in bursting and non-bursting subicular principal cells and interneurons.278 The predominant subicular response to EC stimulation was weak inhibition, suggesting that EC bypasses the hippocampus, modulating the output of the subiculum and thus hippocampal–cortical interaction. Finally, a small depolarising response is observed when CA3 is stimulated and there is no response to dentate gyrus stimulation.263 Commins et al found in vivo paired pulse facilitation (interstimulus interval of 50 ms) as well as LTP by high-frequency stimulation and TBS at the CA1–subiculum synapse.279–281 One hertz low-frequency CA1 stimulation induced frequency-dependent LTD in bursting neurons and LTP in regular spiking subicular neurons, and this bidirectional plasticity relied on the co-activation of muscarinic ACh receptors.282 Finally, EC–subiculum synapses respond to low frequency–induced LTD283 and high frequency–induced LTP.284
In parallel, several studies have reported that subicular neurons show spatially selective firing.80,285–287 Subicular neurons can be divided into three general classes: neurons coding head direction, neurons the firing rate of which reflects position but is modulated by head direction and neurons encoding place.288 The main output of CA1 is subiculum, but subicular place fields appear to be of lower resolution than those of CA1.287 Interestingly, the CA1 and subiculum have been found to operate in a complementary fashion to encode information in a spatial delayed-non-match-to-sample task.289 Subicular neural responses in this task were generally related to shorter delays (15 s or less); conversely, CA1 neural activity was related to long-delay (>15 s) trial-specific information. Finally, the subiculum receives a direct projection from the perirhinal cortex, where neurons are responsive to the novelty or familiarity of objects encountered in the environment but subicular cell firing correlates best not with object novelty or familiarity, but with the concurrent location and speed of animals within the task environment.290
Anterior thalamic nuclei
As a node within the Papez circuit, the ATN gather two main types of neurons: local GABAergic and glutamatergic thalamocortical neurons.291,292 The ATN are composed of the anteroventral, anterodorsal and anteromedial thalamic nuclei. They are reciprocally connected with the presubicular, retrosplenial and cingulate cortices, and their major input arises in the MB. The ATN are distinguished by their subcortical sources of input, which are mainly derived from the hippocampal formation, either directly from the subicular complex257,293,294 or through a relay in MB.295,296 The para/postsubiculum distributes selectively to the anterodorsal thalamic nucleus, the presubiculum to the anteroventral thalamic nucleus and the proximal part of the subiculum almost exclusively to the anteromedial thalamic nucleus.257,267 The hippocampal projection to the MB, which comprises a major part of the descending columns of the fornix, predominantly originates from the dorsal subiculum and to some degree from the presubiculum and parasubiculum.257,294,297
The ATN carry electrophysiological and functional properties related to the intrinsic hippocampal rhythms (tables 3 and 4). Neurons firing rhythmically with theta, the so-called theta cells, and neurons firing selectively when a rat is oriented in a particular direction (41 Hz), the so-called head-direction cells,288,298 coexist in the ATN. Membrane properties of ATN neurons enable them to fire at frequencies between 5 and 11 Hz,299–301 so that some neurons in all divisions of ATN fire rhythmically with theta (6–11 Hz),302–304 with the highest percentage in the anteroventral thalamic nucleus (35%–75%).301–303 In freely moving rats, the great majority of anteroventral thalamic nucleus cells are bursting (87.5%) and 23.7% of those bursting thalamic units are highly entrained to limbic theta rhythm, which corresponds to periods of high locomotor activity. Their bursting is phase-locked to the trough of the local field oscillation.304
Stimulation of the pre-commissural fornix in rodents, along with main findings
Stimulation of the post-commissural ventral fornix in rodents and human patients, along with main findings
There is little overlap between theta rhythm and head-direction neurons. Instead, these cells are mainly present in the anterodorsal nucleus of the thalamus.288,305,306 Such a segregation is also observed in other nodes of the circuit of Papez such as posterior cingulate cortex, the subiculum, the hippocampus and the EC.288,307 Thus, the various subnuclei comprising these parallel, but segregated, theta and head-direction systems are themselves anatomically interconnected, with little crossover between systems. These systems may functionally interact at multiple levels of the circuit to process head-direction information used for spatial learning and navigation.308
Mamillary bodies
The lateral and medial MB are connected with different subregions in the same structures. Dense hippocampal projections from the rostral (septal) subiculum pass through the post-commissural fornix to terminate in the medial MB while parallel projections to the lateral MB arise from the presubiculum, parasubiculum and postsubiculum.309,310 Moreover, whereas the medial MB projects ipsilaterally to the anterior medial and anterior ventral thalamic nuclei, the lateral MB projects bilaterally to the anterior dorsal thalamic nucleus. Finally, the lateral MB have reciprocal connections with the dorsal tegmental nucleus of Gudden, and the medial MB has reciprocal connections with the ventral tegmental nucleus of Gudden. Both lateral and medial MB are also innervated by the supramammillary nuclei, the tuberomammillary nucleus and the septal region.31
Inputs to MB from both the hippocampal formation and the prefrontal cortex are excitatory, but the projections from the tegmental nuclei are inhibitory.310 MB efferents to both anterior thalamic and tegmental nuclei are excitatory.311,312 Neurochemically, the efferents from MB to the ATN use glutamate, aspartate and enkephalin.312
Head-direction neurons are found in the lateral MB, but not the medial MB of the rat. Head-direction signals in the lateral MB precede the signal in the anterior thalamus indicating that the lateral mammillary signal helps to drive the thalamic signal.305,313 Moreover, medial MB neurons fire rhythmically in phase with hippocampal theta,314 whereas few such cells exist in the lateral MB. Septal inactivation eliminates theta activity in the MB but not in the adjacent supramammillary nucleus,48 suggesting that MB is part of a descending system driven from the septum/hippocampus, whereas the supramammillary nucleus is a part of an ascending system generating theta.315 Thus, the MB are a key relay of hippocampal theta rhythm to the ATN and distal circuits.
The MB likely contribute to memory via processes at least in part independent from their hippocampal inputs, such as afferents from the limbic mesencephalon.31 For instance, the pathway responsible for maintaining head direction in rats originates in the dorsal tegmental nucleus of Gudden and projects to the lateral MB, terminating in the anterodorsal thalamic nuclei. Conversely, the regulation of theta rhythm and the optimisation of synaptic plasticity originates in the ventral tegmental nucleus of Gudden, projects to the medial MB and terminates in the anteroventral thalamic nuclei.307,313
Subiculothalamic projections
Anterior thalamic functions rely on direct hippocampal inputs through the direct pathway, as well as indirect information via the indirect pathway.316 The subicular cells projecting to the MB also project to the EC, while the direct subiculothalamic cells do not.271 Since these distinct cell populations may not mediate the same functions, knowing the electrophysiological properties of their synapse with ATN neurons, especially their plasticity and latent periods, is crucial in order to modulate them appropriately with DBS.
Electrical stimulation of the dorsal fornix (figure 2) evokes distinct electrophysiological responses due to stimulation of direct and indirect subiculothalamic cells. In particular, a triphasic response is seen in ATN neurons consisting of a small negative wave followed by a small positive wave and then a long negative wave, with latent periods of 1.5 to 4 ms.317 After a first shock applied to the MB, a period of decreased responsiveness follows the orthodromic activation of AT cells, with a peak at 40 ms and lasting 70–80 ms. MB stimuli delivered at low frequencies ≤1 Hz or >2 Hz evoke respectively a 220 ms long triphasic or monophasic inhibitory postsynaptic potentials in most ATN neurons of preclinical models.291 The hippocampo-mammillary axon terminals were stimulated by Laxton et al at 3 Hz in six patients with AD: the peak of the first evoked response had a 38–52 ms latency and was localised to the hippocampal and parahippocampal gyri, likely corresponding to antidromic activation. At longer latencies (102–256 ms), significant activation of the posterior cingulate gyrus and precuneus area of the parietal lobe was seen suggesting previous activation of the ATN to low frequencies.
Prolonged high-frequency stimulation applied to either the dorsal fornix or the mammillothalamic tract (MTT) did not result in long-lasting thalamic theta activity in rats (table 5). Moreover, 15 min–1 Hz low-frequency stimulation of the dorsal fornix induced augmentation of thalamic low-theta and high-theta over delta ratios for about 120 min in parallel with depressing thalamic synaptic responses whereas the same protocol applied to MTT failed to evoke significant oscillatory changes.301,304 Synaptic depression has been proposed as a dynamic gain control mechanism in cortical information processing318 and hippocampal theta may modify thalamic responsiveness to stimuli coming from the tegmental area via the MB. Differences in basal synaptic transmission, short-term and long-term synaptic plasticity were found between the hippocampo-thalamic and mammillothalamic tracts. A brain-derived neurotrophic factor-dependent augmentation of synaptic transmission was observed only at mammillothalamic synapses. Paired-pulse stimulation, however, induced facilitation in both pathways. The amplitude of the thalamic activity was readily potentiated after high-frequency stimulation of the mammillothalamic tract but not of the dorsal fornix. Low-frequency stimulation of the mammillothalamic tract induced potentiation.301,304 Seemingly, the two major inputs to the ATN have opposing or complementary actions.
Stimulation of the dorsal fornix or the mammillothalamic tract in rodent models
Following clinical trials of human fornix stimulation, Hescham showed in 2016 that 1-hour stimulation at 100 Hz of the hippocampo-mammillary axon terminals induced a selective activation of cells in the CA1 and CA3 subfields of the rodent dorsal hippocampus.319 In addition, they observed a substantial increase in the levels of extracellular hippocampal ACh, which peaked 20 min after stimulus onset, whereas hippocampal glutamate levels did not change compared with baseline. In a rat model of scopolamine-induced dementia, acute bilateral DBS of the hippocampo-mammillary axon terminals reversed the memory impairing effects of scopolamine in the object location task.320 Both 10 Hz and 100 Hz stimulations were found to be efficient, but a higher current density threshold was needed at 10 Hz.
Human forniceal subiculo-mammillary stimulation
Several trials have documented the clinical effects of chronic forniceal subiculo-mammillary axon terminals for DBS (table 4). In a patient with morbid obesity, bilateral DBS to the hypothalamus, which is closely associated with the ventral post-commissural fornix, elicited recall of autobiographical memories.3 Subsequently, an open-label phase I trial of fornix DBS was initiated: patients with mild-to-moderate AD were implanted with electrodes 2 mm anterior to the columns of the ventral post-commissural fornix. Patients received high-frequency DBS for 12 months, and PET studies a year later revealed increases in cortical glucose metabolism that were correlated with improved cognitive measures in two orthogonal networks: a frontal-temporal-parietal-striatal-thalamic network and a frontal-temporal-parietal-occipital-hippocampal network.321 The finding of increased glucose metabolism is a striking contrast to the longitudinal metabolic decline generally seen in patients with AD.322,323 Moreover, clinical evaluation of the AD Assessment Scale cognitive subscale (ADAS-Cog) and the Mini Mental State Examination (MMSE) suggested possible slowing in the rate of progressive cognitive decline in certain patients with AD.321,324 In the aforementioned cortical regions, higher baseline metabolism prior to DBS and increased metabolism after 1 year of DBS were correlated with better outcomes in global cognition, memory and quality of life. A single case report showed evidence of stabilisation of MMSE and ADAS-Cog scores at 1-year follow-up and subjective improvement.325 Although a subsequent crossover randomised phase II clinical trial did not yield cognitive benefits in all patients with AD, those over the age of 65 showed a slower decline.326 A multicentre phase III trial is now underway to assess which AD patient subgroup will benefit most from forniceal DBS.5
Subiculothalamic pathway summary
Overall, a characteristic feature of the anterior thalamic neurons is their ability to fire rhythmically in the theta range.327 These 5–12 Hz oscillations in the anterior ventral nucleus of the thalamus receive descending inputs from the subiculum and ascending inputs from the medial MB via the forniceal subiculothalamic pathway.307 Theta rhythm is thought to play a critical role in the mnemonic functions of the limbic system64,315,328 and oscillatory patterns in the theta range may enable synaptic plasticity.109 Furthermore, inactivation of the MS and thus of the forniceal septohippocampal pathway abolishes theta discharge in both the hippocampus and MB,48 ,329 two of the major regions providing inputs to anterior thalamus. Hence, the ATN appear to be part of a descending system driven from the MS via the forniceal septohippocampal pathway, and theta oscillations in the anterior thalamus might complement hippocampal–diencephalic memory processing after propagation through the forniceal subiculothalamic pathway.308
Overall summary and future directions
Understanding the complex interconnections within the circuit of Papez mediated by the fornix sheds light on memory function in healthy and disease states. This framework is also relevant in the context of the design of future clinical forniceal DBS strategies. The forniceal septohippocampal axis generates brain oscillations that are central to memory processes, such as theta and gamma oscillations, as well as theta–gamma cross-frequency coupling (table 6). The forniceal subiculothalamic pathway relays these rhythms across the nodes of the circuit of Papez, mediating diverse functional aspects of memory. The topography of projections and interplay of intrinsic rhythms give rise to synaptic plasticity and memory consolidation. Given the complex neurophysiology and connectivity of subnodes within the circuit of Papez, future neuromodulation devices should strive to deliver individualised therapy in various nodes of the circuit of Papez in response to real-time electrophysiological data.330 Biotechnological innovations in the field of neuromodulation are needed to optimise sensing and delivery algorithms, as well as power management strategies in future DBS devices. Prospective randomised and double-blinded human trials are underway to evaluate the true potential of DBS to rescue memory deficits in patients with neurodegenerative, vascular or traumatic lesions of the circuit of Papez.5 Please see online supplementary file 1 for references 61 to 308.
Supplemental material
Presence of theta and/or gamma oscillations within the various nodes of the Papez circuit interconnected by the fornix, and effects of electrical stimulation of these nodes
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
SS and AF are joint first authors.
SS and AF contributed equally.
Contributors SS and AF are joint first authors and contributed equally to the manuscript. SS conceived the manuscript and wrote the first draft. AF created figures, wrote the anatomy and pathology chapters and revised the manuscript. EG contributed the imaging and clinical trials chapter. AL oversaw revisions and approved the final manuscript.
Funding This review was made possible by the R.R. Tasker Chair in Functional Neurosurgery, University Health Network (AML) and the University of Manitoba Clinician Investigator Program (AF)
Competing interests AML is a consultant to Medtronic, St Jude, Boston Scientific, Functional Neuromodulation and Insightec.
Patient consent for publication Not required.
Provenance and peer review Not commissioned; externally peer reviewed.