Article Text

Abstract
Objective To investigate in-vivo cortical gyrification patterns measured by the local gyrification index (lGI) in presymptomatic c9orf72 expansion carriers compared with healthy controls, and investigate relationships between lGI and cortical thickness, an established morphometric measure of neurodegeneration.
Methods We assessed cortical gyrification and thickness patterns in a cohort of 15 presymptomatic c9orf72 expansion carriers (age 43.7 ± 10.2 years, 9 females) compared with 67 (age 42.4 ± 12.4 years, 36 females) age and sex matched healthy controls using the dedicated Freesurfer pipeline.
Results Compared with controls, presymptomatic carriers showed significantly lower lGI in left frontal and right parieto-occipital regions. Interestingly, those areas with abnormal gyrification in presymptomatic carriers showed no concomitant cortical thickness abnormality. Overall, for both presymptomatic carriers and healthy controls, gyrification and cortical thickness measures were not correlated, suggesting that gyrification captures a feature distinct from cortical thickness.
Conclusions Presymptomatic c9orf72 expansion carriers show regions of abnormally low gyrification as early as their 30s, decades before expected symptom onset. Cortical gyrification represents a novel grey matter metric distinctive from grey matter thickness or volume and detects differences in presymptomatic carriers at an early age.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
A hexanucleotide expansion in c9orf72 (C9) is the most common genetic cause of familial and sporadic frontotemporal dementia (FTD) and amyotrophic lateral sclerosis (ALS).1 2 Previous studies show that presymptomatic c9orf72 expansion carriers (preSxC9) have deficits in grey matter volume, white matter integrity3–5 and disruption of functional connectivity networks as soon as their early 30s, decades before expected symptom onset.4 These abnormalities may represent incipient neurodegeneration, abnormal brain development or both. Whether such deficits represent neurodevelopmental differences vs early atrophy, however, remains an open question.
Brain development is an intricate and complex process that begins during gestation and continues through early adulthood. Cortical folding, or gyrification, represents a dynamic phenomenon allowing for abundant expansion of the cortical surface.6 7 During the third trimester of fetal life, gyrification proliferates rapidly, and the brain grows from a relatively smooth surface towards a gyrification pattern resembling the adult brain.8 9 Gyrification declines exponentially throughout childhood and adolescence, and then declines more linearly during adulthood, and these declines in gyrification arise in parallel with age-related volume loss.7 10 With novel morphometric MRI techniques, three-dimensional cortical folding patterns can be evaluated in-vivo by computing the local gyrification index (lGI), defined as the ratio between the cortex buried within sulcal folds and the area of the cortical surface.11
In children with neurodevelopmental disorders such as dyslexia and autism, specific brain regions show a higher degree of cortical folding compared with typically developing children.12–15 In dyslexia, this abnormally higher gyrification may be due to the absence of the age-related gyrification decline seen in typically developing children. Studies in adults have revealed abnormal gyrification trajectories in patients with major psychiatric disorders (ie, depression, bipolar disorder and schizophrenia)10 and neurodegenerative disorders such as Parkinson’s and Alzheimer’s diseases.16 17 In contrast to the pattern seen in neurodevelopmental disorders, patients with Alzheimer’s disease show abnormally low gyrification associated with sulcal widening, reduced cortical thickness and cognitive decline.16
To date, no study has applied this novel metric to assess gyrification in presymptomatic carriers of any neurodegenerative disease-causing mutations. We hypothesised that presymptomatic c9orf72 expansion carriers would have abnormally increased cortical gyrification as seen in dyslexia and autism, reflecting neurodevelopmental differences. In the present study, we assessed cortical gyrification and cortical thickness in 15 preSxC9 compared with age and sex matched healthy controls (HC).
Methods
Subjects
We compared 15 preSxC9 (age 43.7±10.2 years, 9 females) to 67 (age 42.4±12.4 years, 36 females) age and sex matched HC. Demographic characteristics and neuropsychological test scores did not statistically differ between groups and are reported in a previous study.4 In genetic FTD, mean familial age of onset correlates with age of symptom onset in symptomatic carriers.5 Thus, the estimated time to symptom onset for a presymptomatic carrier was calculated by determining the difference between the carrier’s age and the mean familial age of onset. The estimated time to symptom onset of the presymptomatic carriers in the present study was 8.2±11.0 years. The University of California, San Francisco Committee on Human Research approved the study. Participants provided informed consent prior to participation.
Image acquisition
All subjects underwent MRI scanning on a Siemens Tim Trio 3T scanner. Structural volumetric T1-weighted imaging was acquired with the following parameters: TR/TE 2300/2.98 ms; flip angle 9; 160 sagittal slices; voxel size=1 mm3; matrix size 240 x 256.4
Freesurfer-based cortical thickness and lGI analyses
The preprocessing of T1-weighted images was performed using FreeSurfer V.5.3 (http://surfer.nmr.mgh.harvard.edu/) in accordance with a standard autoreconstruction algorithm. The pipeline included intensity normalisation, removal of non-brain tissue using a watershed algorithm technique, generation of the surface mesh representations using a surface deformation algorithm, tessellation of the grey matter boundary, automated topology correction and Talairach-Tournoux transformation.18–20 The accuracy of the segmentation results was validated by a neuroradiologist (EC). Cortical thickness measures were obtained based on the shortest distance between the grey matter/ white matter and grey matter/ cerebrospinal fluid boundaries at each vertex of the tessellated surface. Gyrification of the entire cortex was also assessed using FreeSurfer. lGI was computed using the method described by Schaer and colleagues.11 21 Cortical thickness and lGI images were normalised to the fsaverage space (MNI305 template) and smoothed with a full-width at half maximum Gaussian kernel of 15 mm for cortical thickness and 5 mm for lGI (figure 1).7

Example of an individual subject lGI map. (A) Lateral view of the native space cortical surface of one healthy control subject (male, 57 years old) and (B) lateral view of the lGI map overlaid on the FreeSurfer fsaveragetemplate. Regions with a higher lGI indicate a greater degree of gyrification. The ‘heat’ colour bar represents lGI values, ranging from 0 to 5. lGI, local gyrification index.
Statistical analysis with AFNI
We examined group differences (preSxC9 vs HC) in cortical thickness and lGI using independent two-sample t-tests (AFNI 3dttest++, www.afni.nimh.nih.gov) at two thresholds: p<0.001 uncorrected and p<0.05 corrected for family-wise error (FWE). We performed the FWE multiple comparison correction using the ‘slow_surf_clustsim.py’ and ‘quick.alpha.vals.py’ tools in the AFNI software package. We chose to report results at p<0.001 uncorrected, consistent with the previous study.4 These t-tests perform a vertex-by-vertex regression over the whole cortical surface to estimate the mean difference of the input data between the two groups. Participants’ age, gender and cortical thickness (for the lGI comparison) were included as covariates of no interest.22 To explore potential differences between groups with respect to age, we examined group as an interaction term for the relationship between lGI and age. We also computed the effect size measured by Cohen’s d (d) for each cluster found to have significantly lower lGI in the presymptomatic carriers; this was obtained by dividing the difference between mean values by the common SD of the two groups.23 With the effect size, we performed a power calculation for a two-tailed analysis using an alpha value of 0.05.
Relationship between cortical thickness and lGI
We performed a Pearson correlation analysis across the grey matter in all subjects to assess for potential associations between cortical thickness and lGI. We also performed a least squares regression (LSR) analysis to quantify the variance attributable to group, age or cortical thickness within the map of regions in which preSxC9 showed significantly abnormal lGI compared with HC. We used the JMP PRO 14 statistical programme to perform this analysis.
Results
PreSxC9 show regions of lower gyrification compared to healthy controls
PreSxC9 showed four regions of lower lGI compared with controls (pFWE <0.05) in the left subgenual anterior cingulate cortex, left precentral gyrus, right inferior parietal lobule and right superior occipital gyrus (figure 2 and table 1). The effect size measured by Cohen’s d was large (d=1.0) in each significant cluster with a power of 93.3%. A trend (p<0.001 uncorrected) towards lower lGI emerged in the left orbitofrontal, insular and occipital cortices and right parietal cortex (figure 2 and table 1). There were no regions for which preSxC9 had a significantly greater lGI compared with controls.

PreSxC9 show regions with low local gyrification index. Regions with significantly lower local gyrification index emerged in preSxC9 compared with controls (p<0.001 uncorrected, cyan). Regions in left precentral gyrus, left subgenual anterior cingulate, right superior occipital cortex and right inferior parietal lobule remained significant at pFWE <0.05 (dark blue). FWE, family-wise error.
Regions showing lower local gyrification in preSxC9
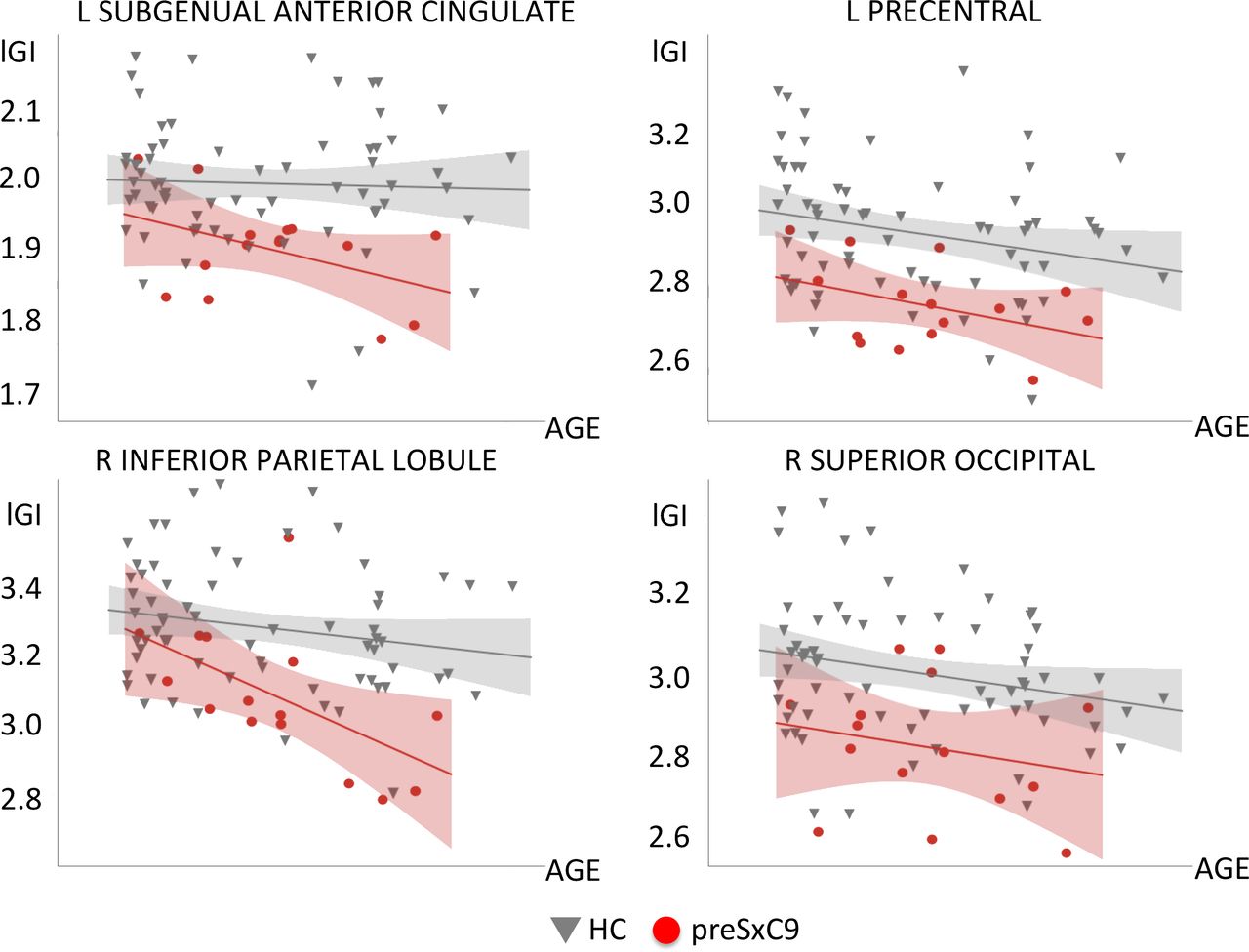
We next explored whether carriers and controls might have different cross-sectional relationships between gyrification and increasing age. Across the entire cortical surface, there were no statistically significant regions for which the slope of lGI vs age differed between carriers and controls at either threshold. Within those regions showing lower lGI for preSxC9, both groups showed lGI declines with age, and the slopes of these lGI declines with age did not significantly differ between groups (figure 3).

{kind=link}
{kind=link}
{kind=link}
Relationships with age for regions showing low lGI. We extracted the mean lGI within each of the four regions showing low lGI in preSxC9 compared with HC (at pFWE <0.05). These clusters appeared within left precentral gyrus, left subgenual anterior cingulate, right superior occipital gyrus and right inferior parietal lobule. HC (dark grey), preSxC9 (red) fit lines are shown for visualisation purposes only. Age axis labels are removed to protect participant anonymity. FWE, family-wise error; HC, healthy controls; lGI, local gyrification index.
Compared with controls, preSxC9 did not show any differences in cortical thickness at pFWE <0.05. Regions of lower cortical thickness in preSxC9 emerged at p<0.001 uncorrected in frontal, insular, cingulate and occipital cortex bilaterally (online supplementary data). There were no regions for which preSxC9 had a significantly greater cortical thickness compared with controls at p<0.001 uncorrected.
Supplemental material
Gyrification and cortical thickness are uncorrelated
To probe whether lGI represents a metric independent from cortical thickness, we correlated cortical thickness and lGI for the entire grey matter across all subjects. There was no significant correlation between cortical thickness and lGI at pFWE <0.05. We next correlated lGI and thickness within each subject subgroup, and no correlations emerged at either threshold for the preSxC9 or the HC groups. Overall, these analyses suggest that lGI and thickness capture independent characteristics of grey matter within our study cohort.
To quantify the relative contributions of group, cortical thickness, age and sex to the variance of lGI, we performed an LSR analysis for regions in the preSxC9 <HC lGI map (pFWE <0.05). The model that best explained lGI variability (R2=0.37; p<0.0001) included group (beta=−5.30; p<0.0001) and age (beta=−3.26; p=0.002). As expected, the variance within regions of low gyrification in preSxC9 was most strongly attributable to group, then age. Interestingly, cortical thickness and sex were not predictors of lGI.
Discussion
PreSxC9 feature structural brain deficits decades before expected symptom onset, with grey and white matter declines with age at a rate similar to controls,4 leading to the intriguing hypothesis that such deficits may represent neurodevelopmental differences. Because neurodevelopmental disorders such as dyslexia and autism feature regions of abnormally increased gyrification, we had hypothesised that preSxC9 would also show cortical regions of ‘hypergyrification’. In contrast to our hypothesis, we found that preSxC9 had no regions of increased gyrification, but instead regions of low lGI in the left subgenual anterior cingulate cortex, left precentral gyrus, right inferior parietal lobule and right superior occipital gyrus. Both preSxC9 and controls showed decreasing lGI across the adult age span studied, without any difference in slopes for the relationship between lGI and age. Low lGI values appeared in carriers as young as their 30s, consistent with previous findings that preSxC9 have grey matter deficits at this age.4 Interestingly, regions of low gyrification in preSxC9 only partially overlapped with regions of low grey matter thickness and volume, and within regions of low gyrification, cortical thickness was not a significant predictor of lGI variance. Moreover, gyrification and thickness were uncorrelated across the entire cortex. These findings support the notion that gyrification captures a feature of the cortex that is distinctive from grey matter thickness or volume and that gyrification may prove a useful marker for detecting differences in presymptomatic carriers.
PreSxC9 have low gyrification in regions atrophied during the symptomatic phase
As with sporadic bvFTD, patients with bvFTD due to the c9orf72 expansion show symmetric, prominent atrophy in the anterior insula, anterior cingulate and frontotemporal cortex.24–27 In contrast to sporadic bvFTD, c9orf72-bvFTD features parietal, occipital26 27 and thalamic atrophy.28 Similarly, patients with ALS due to the c9orf72 expansion show a more distributed atrophy pattern, with a greater degree of frontal and anterior cingulate atrophy29 and thalamic degeneration30 compared with sporadic ALS. PreSxC9 show grey matter deficits in regions that atrophy in c9orf72-bvFTD, including the anterior cingulate, medial frontal and dorsolateral prefrontal cortex, insula, precentral gyrus, precuneus, occipital cortex, striatum and medial thalamus.4
The regions with low gyrification in preSxC9 are congruent with regions of grey matter atrophy in c9orf72-bvFTD28 and grey matter volume deficits in preSxC9.4 PreSxC9 showed low lGI in the anterior cingulate cortex, which represents a key region of atrophy in bvFTD with or without c9orf72.28 31 The low lGI found in the primary motor cortex in presymptomatic carriers may presage future vulnerability to motor neuron disease that manifests during the symptomatic phase. Regions of low lGI also emerged in parieto-occipital cortex, consistent with grey matter atrophy and deficits in c9orf72-bvFTD and presymptomatic carriers.4 28
Low gyrification in preSxC9: neurodevelopmental versus neurodegenerative?
Whether regions of low gyrification in preSxC9 represent a neurodevelopmental difference or early atrophy remains an open question. We identified regions of low gyrification in carriers as young as their 30s, presumably decades before expected symptom onset. This finding is parallel to a previous study in which these same presymptomatic carriers showed grey matter volume and white matter deficits at a similar age.4 In the present study and the previous study, both lGI and grey and white matter declined with age in preSxC9 at rate similar to controls, suggesting that these deficits may be due to neurodevelopmental differences.
Previous studies have revealed that children with dyslexia and autism show regions of abnormally high lGI. In dyslexia, regions with abnormally increased lGI correlate with neurite morphology, possibly associated with synaptic pruning deficits.15 In contrast to these developmental disorders in children, preSxC9 showed low lGI compared with controls. One possibility is that low gyrification represents a developmental deficit in cortical expansion in preSxC9, resulting in low lGI throughout the entire lifespan. A second possibility is that low lGI values could reflect that preSxC9 subjects ‘overprune’ gyri during adolescence and early adulthood to a degree beyond the typical decline in gyrification seen during adulthood. If due to developmental differences, these regions with low lGI may represent vulnerable regions that set the path for focal future neurodegeneration. A third consideration is that low lGI could represent early atrophy in preSxC9. PreSxC9 show gyrification declines with age at a rate similar to controls. If low lGI were to represent atrophy, however, we would expect to see accelerated lGI decline as seen in Alzheimer’s and Parkinson’s diseases,16 17 yet our cross-sectional data did not show an accelerated decline in lGI for older carriers. Future longitudinal studies, including those with children and adolescents, are needed to establish trajectories of lGI and other imaging metrics to determine the natural history of brain development.
Relationships between gyrification and other imaging measures
Our data suggest that the regions of low gyrification in preSxC9 were not attributable to cortical thickness deficits, consistent with other studies that have examined the relationship between lGI and other grey matter metrics. In a study of typically developing children and healthy adults, lGI correlated with grey matter volume but not with cortical thickness.7 In parallel, a study of adolescents with autism also showed that lGI and cortical thickness were uncorrelated, and that instead, lGI correlates with white matter architecture and connectivity.32
Previous studies suggest that gyrification has stronger associations with structural white matter connectivity rather than cortical thickness. In dyslexia, abnormally increased gyrification is correlated with neurite architecture changes as measured by neurite orientation dispersion and density imaging15; in autism spectrum disorder, increased gyrification is associated with altered white matter organisation.12 Although the mechanics of gyrification remain opaque, one theory suggests that cortical folding is driven to optimise white matter connections, as tension-mediated forces along axons bring highly connected regions closer to each other.6 In support of this theory, the immense increase in brain gyrification during the late fetal period coincides with an explosive development of cortico-cortical connections.6 15 33–36 Future studies will inform the relationships between white matter structural connectivity and gyrification in preSxC9 and other neurodevelopmental disorders.
Conclusions
PreSxC9 show regions of abnormally low gyrification in regions known to be targeted in c9orf72-FTD and c9orf72-ALS. If neurodevelopmental, deficits in gyrification and grey matter volume may confer vulnerability that predisposes selective brain regions to future neurodegeneration. Cortical gyrification represents a novel grey matter metric, and future longitudinal studies will clarify if low gyrification in preSxC9 represents a neurodevelopmental phenomenon.
Acknowledgments
We thank all participants and their families whose help and participation made this work possible.
References
Footnotes
EC and GB contributed equally.
MLG-T and SEL contributed equally.
Contributors EC and GB designed and conceptualised the study. They processed the data,
performed the statistical analysis of the data and drafted the manuscript for intellectual content. SAC, AMK and WS were involved in the data collection. HLR, TPZ, GC, DHG and RR were involved in the data collection and revision of the manuscript for intellectual content. BLM revised the manuscript for intellectual content. MLG-T and SEL were involved in the design and conceptualisation of the study and revision of the manuscript for intellectual content.
Funding This work was supported by the National Institutes of Health [SEL: R01 AG058233, K23AG039414; TPZ: F32AG030249,R01MH096861; MGT: R01NS050915, K24DC015544; GC: AG035610; RR: R35NS097261; BLM:P01AG019724, P50AG23501]. The John Douglas French Alzheimer's Foundation[GC]. State of California DHS04-35516[MGT]. Samples from the National Cell Repository for Alzheimer's Disease(NCRAD), which receives government support under a cooperative agreement grant(U24AG21886) awarded by the National Institute on Aging (NIA), were used in this study.
Competing interests None declared.
Patient consent for publication Not required.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Researchers may obtain imaging code used for preprocessing and statistical analysis in this study from the corresponding author on reasonable request. All data relevant to the study are included in the article or uploaded as supplementary information.