Article Text

Abstract
Objective: To examine the properties and potential clinical uses of myogenic potentials to bone conducted sound.
Methods: Myogenic potentials were recorded from normal volunteers, using bone conducted tone bursts of 7 ms duration and 250–2000 Hz frequencies delivered over the mastoid processes by a B 71 clinical bone vibrator. Biphasic positive–negative (p1n1) responses were recorded from both sternocleidomastoid (SCM) muscles using averaged unrectified EMG. The best location for stimulus delivery, optimum stimulus frequency, stimulus thresholds, and the effect of aging on evoked response amplitudes and thresholds were systematically examined. Subjects with specific lesions were studied. Vestibular evoked myogenic potentials (VEMP) to air conducted 0.1 ms clicks, 7 ms/250–2000 Hz tones, and forehead taps were measured for comparison.
Results: Bone conducted sound evoked short latency p1n1 responses in both SCM muscles. Ipsilateral responses occurred earlier and were usually larger. Mean (SD) p1 and n1 latencies were 13.6 (1.8) and 22.3 (1.2) ms ipsilaterally and 14.9 (2.1) and 23.7 (2.7) ms contralaterally. Stimuli of 250 Hz delivered over the mastoid process, posterosuperior to the external acoustic meatus, yielded the largest amplitude responses. Like VEMP in response to air conducted clicks and tones, p1n1 responses were absent ipsilaterally in subjects with selective vestibular neurectomy and preserved in those with severe sensorineural hearing loss. However, p1n1 responses were preserved in conductive hearing loss, whereas VEMP to air conducted sound were abolished or attenuated. Bone conducted response thresholds were 97.5 (3.9) dB SPL/30.5 dB HL, significantly lower than thresholds to air conducted clicks (131.7 (4.9) dB SPL/86.7 dB HL) and tones (114.0 (5.3) dB SPL/106 dB HL).
Conclusions: Bone conducted sound evokes p1n1 responses (bone conducted VEMP) which are a useful measure of vestibular function, especially in the presence of conductive hearing loss. For a given perceptual intensity, bone conducted sound activates the vestibular apparatus more effectively than air conducted sound.
- tone burst
- bone conduction
- vestibular reflexes
- AC, air conducted, BC, bone conducted
- EMG, electromyography
- HL, hearing level
- NHL, normal hearing level
- SCM, sternocleidomastoid muscle
- SPL, sound pressure level
- VEMP, vestibular evoked myogenic potentials
- VOR, vestibulo-ocular reflexes
Statistics from Altmetric.com
- AC, air conducted, BC, bone conducted
- EMG, electromyography
- HL, hearing level
- NHL, normal hearing level
- SCM, sternocleidomastoid muscle
- SPL, sound pressure level
- VEMP, vestibular evoked myogenic potentials
- VOR, vestibulo-ocular reflexes
The assessment of patients with vertigo and disequilibrium can be challenging, and the availability of investigations that examine each component of the vestibular apparatus would facilitate a more complete evaluation. Vestibulo-ocular reflexes (VOR) to caloric stimulation and sinusoidal rotation have long been in clinical use as measures of lateral semicircular canal function. More recently, impulsive tests which measure VOR to rapid head accelerations in the plane of each semicircular canal have allowed measurement of individual canal function.1 Clinically useful techniques that assess otolith function, however, are few and not widely available.2 The click evoked myogenic potential—a relatively recently developed simple, non-invasive test of otolith function—is based on the acoustic sensitivity of the saccule.3
Intense clicks delivered through headphones evoke vestibular dependent, biphasic potentials (p13n23) recordable from the sternocleidomastoid (SCM) muscles ipsilateral to the stimulated ear, during tonic activation (vestibular evoked myogenic potentials (VEMP): Colebatch et al, 19943). Their short onset latency of about 8 ms suggests that these responses are mediated by an oligosynaptic arc, which probably consists of the primary vestibular afferents synapsing at the vestibular nuclei, the second order vestibulocollic neurones, and the neck motor neurones innervating the SCM. Intense short tone bursts delivered through headphones also evoke VEMP which have similar properties to those evoked by clicks when stimuli of appropriate frequency and duration are used.4,5 Both click evoked and tone burst evoked responses require the integrity of middle ear conductive mechanism to convey the stimulus to the vestibular end organs.3,6 To enable measurement of VEMP in subjects with conductive hearing loss, Halmagyi et al proposed the use of forehead taps delivered through a modified reflex hammer, which probably results in a wave of vibration that is directly conveyed to both labyrinths, circumventing the middle ear conductive apparatus.7 This stimulus evokes an initial positivity (p1) followed by two negativities (“n1,n2,” see fig 1), of which only the first is vestibular dependent.7 The precise vestibular afferents stimulated by a forehead tap have not been established, although an otolith contribution is likely, given the similarity of the waveform to that of the click evoked response. Sheykholeslami et al recently reported that tone bursts delivered by a clinical bone vibrator applied over the mastoid process give rise to short latency potentials in the SCM, which were likely to be of vestibular origin.8 This new technique holds promise as an alternative to the tap evoked response, as a method of eliciting vestibular dependent responses in subjects with conductive hearing loss. It has the advantage of being a calibrated and reproducible stimulus and therefore should elicit consistent responses and enable the measurement of stimulus thresholds.
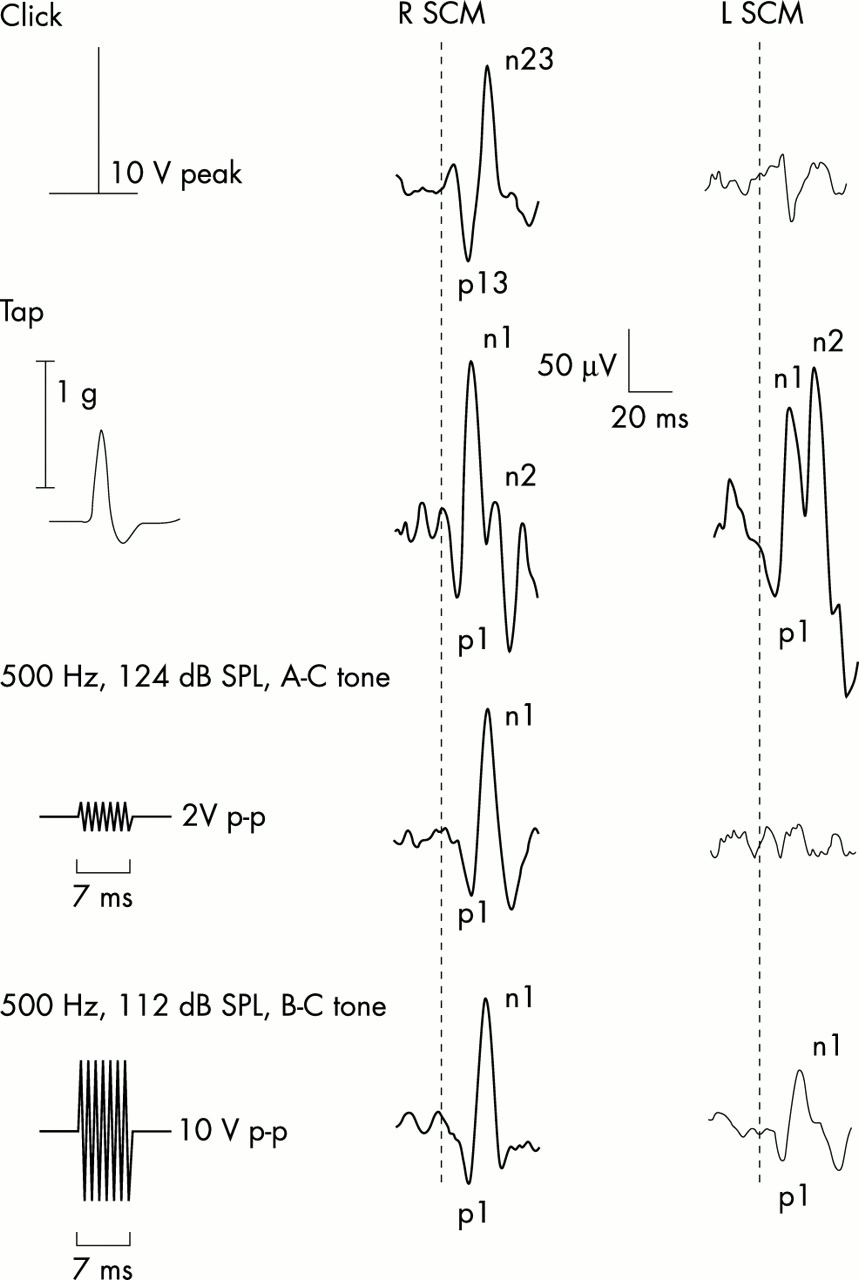
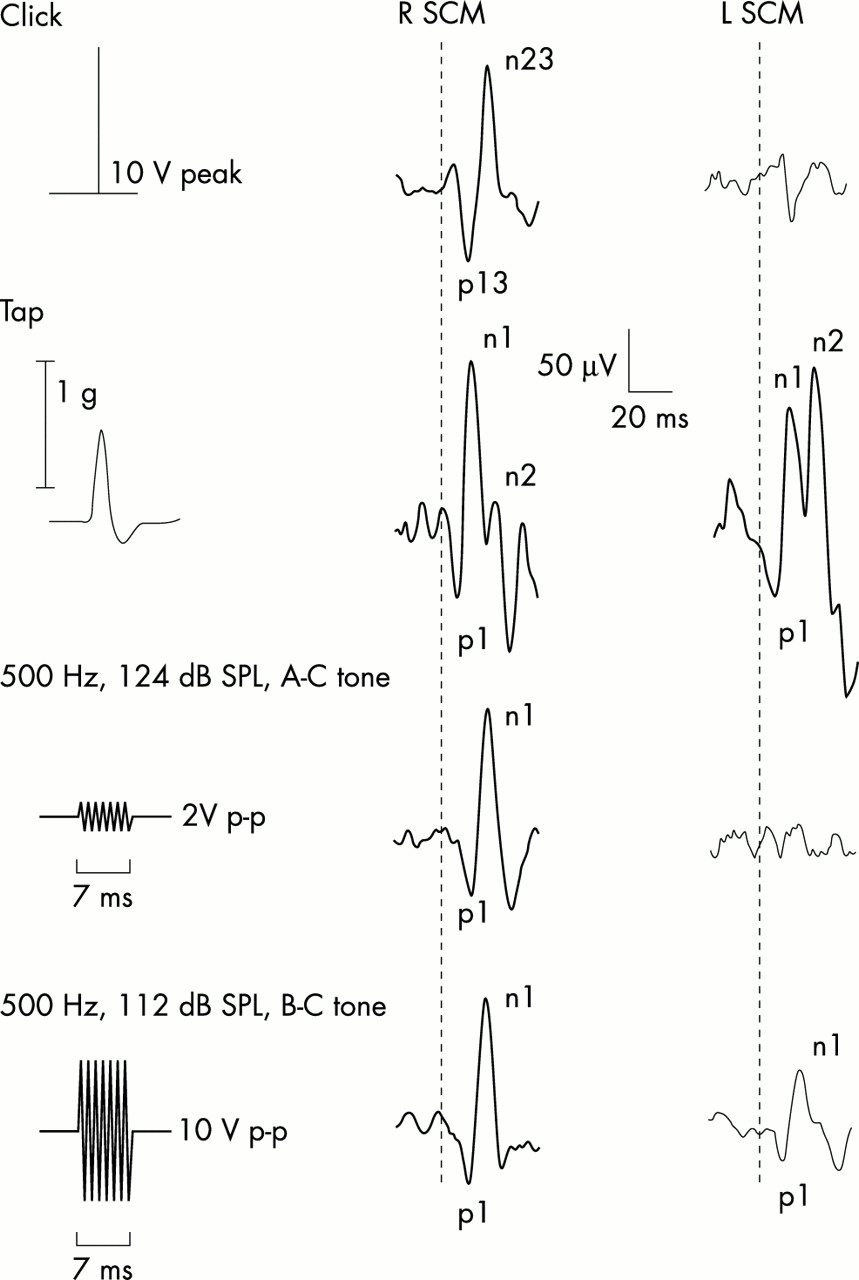
Myogenic potentials to four stimuli in a 25 year old subject. The traces consist of averaged unrectified electromyographic recordings from the left and right sternocleidomastoid muscles (SCM). The thick and thin traces represent responses ipsilateral to and contralateral to the side of stimulation, respectively. 100 dB NHL clicks delivered in the right ear evoked p13n23 responses in the ipsilateral SCM. Forehead taps produced bilateral symmetrical vestibular evoked myogenic potentials (VEMP) by activating both labyrinths; these consisted of an initial positivity followed by two negativities (n1,n2), of which the first is vestibular dependent. Air conducted (AC) tones (124 dB SPL/500 Hz/7 ms), like clicks, give rise only to ipsilateral p1n1 responses. Bone conducted (BC) tones (112 dB SPL/500 Hz/7 ms) result in p1n1 responses bilaterally; the ipsilateral response being larger and occurring earlier in most subjects. Corrected reflex amplitudes for clicks, taps (averaged for left and right sides), air conducted tones, and bone conducted tones were 1.5, 2.1, 1.64, and 2.05, respectively. Tone amplitudes are plotted to scale; head acceleration is shown for taps.
The emergence of several related methods of evoking vestibulocollic reflexes necessitates assessment of their relative merits and definition of the place of each these techniques in vestibular function testing. Our aim in the present study was to examine further the properties of responses evoked by bone conducted sound. We confirmed the vestibular dependence and defined the optimum stimulus parameters, methods of delivery, and reproducibility of bone conducted responses, and compared them with VEMP evoked by air conducted tones, clicks, and taps in the same subjects.
METHODS
Ten healthy adults aged 24 to 52 years (five men, five women) were studied after obtaining informed consent and local ethics committee approval.
Myogenic potentials to 7 ms/500 Hz/112 dB SPL (sound pressure level) tone bursts delivered by a clinical bone vibrator (B71, Radioear Corporation, New Eagle, Pennsylvania, USA) placed on the mastoid process (3 cm posterior to the external acoustic meatus) were recorded from all subjects. VEMP to clicks of 0.1 ms/145 dB SPL (= 100 dB normal hearing level (NHL)) and 7 ms/500 Hz tone bursts of 123.5 dB SPL delivered through headphones (our “standard parameters” for these stimuli) and forehead taps were also measured for comparison. Although values for air and bone conducted sound are not directly comparable, we have expressed all intensities in dB SPL.
Subjects were studied lying on a couch with an adjustable backrest, at 45° from the horizontal plane, while raising their heads against gravity to activate both SCM muscles. Electromyographic (EMG) activity was recorded from surface electrodes over symmetrical sites on the SCM muscles. Active recording electrodes were placed on the junction of the upper and middle thirds of the muscle belly, and reference electrodes on the medial portion of the clavicles. Both rectified and unrectified EMG signals were amplified and bandpass filtered (8 Hz to 1.6 kHz); EMG was sampled (5 kHz) from 20 ms before to 100 ms after the stimulus onset (unless otherwise specified) using a 1401 Plus analogue to digital converter and Sigavg software (Cambridge Electronic Design, Cambridge, UK) on a PC. Evoked responses were measured using unrectified averaged EMG; responses to 256 stimuli were averaged for each ear or side tested. The response amplitudes were measured peak to peak and divided by the mean rectified EMG activity level for the 20 ms preceding the stimulus onset to correct for background activation (“corrected amplitudes”). For VEMP, the relation between the reflex amplitude and tonic EMG activity is linear (Colebatch et al, 19943), and this correction would be expected to control for differences in activation. In the present study, results are presented as both raw (uncorrected) amplitudes and corrected reflex amplitudes. Side to side differences in corrected amplitudes were expressed as an asymmetry ratio (AR); where Al and As were the larger and the smaller amplitudes of the left and right sides,

Myogenic potentials evoked by bone conducted sound
Tone bursts were generated using a Jupiter 2010 function generator (Black Star, Cambridge, UK); these were amplified and gated electronically. The onset and offset of the gate were fixed and were independent of the phase of the input. Bone conducted stimuli were delivered using a B71 clinical bone vibrator previously calibrated using a Bruel and Kjaer (model BMK 4930) artificial mastoid. An initial study was undertaken using stimuli of 7 ms duration/500 Hz/112 dB SPL (“standard parameters”) delivered at three per second; these were based upon the optimum parameters for air conducted tone bursts.5
The effects of varying different parameters (location, tonic activation, frequency) were systematically investigated. Using the standard parameters, myogenic responses were recorded in five subjects with the bone vibrator placed on Fpz, the anterior temporal region (approximately 2 cm above and 4 cm anterior to the external acoustic meatus), the mastoid processes (3 cm posterior to the external acoustic meatus), and the external occipital protuberance. The best location for delivery of the stimulus was mapped in greater detail in 10 subjects, using a grid consisting of 12 points around the external acoustic meatus.
To determine whether bone conducted tone burst evoked response amplitudes also increase linearly with EMG activity, responses to standard stimuli were measured in five subjects, while maintaining different degrees of background activation (approximately 20, 40, 60, 80, and 100 μV). Peak to peak amplitudes were plotted against mean rectified EMG activity. In addition, one subject was studied at 15 different levels of activation corresponding to mean rectified EMG levels of 15 μV to 150 μV. Myogenic potentials were measured in two separate experiments done seven to 10 days apart in 10 subjects to assess between-experiment reproducibility. Reproducibility of tap evoked responses was also examined for comparison.
To determine the optimum stimulus frequency, responses to 250, 500, 1000, and 2000 Hz bone conducted tone bursts were measured in 10 subjects. Stimuli of nearly constant intensities were used (that is, 107.5, 107, 107.5, and 105.5 dB SPL, corresponding to 20, 5, 10, and 10 volts peak to peak respectively at 250, 500, 1000, and 2000 Hz). These stimuli, generated digitally, were phase locked at 0° and had 1 ms rise and fall times and a 7 ms hold time. In each subject, the optimum stimulus frequency was determined and stimulus thresholds were measured at this frequency using 6 dB decrements in intensity.
We have previously reported clear effects of age on VEMP amplitudes and thresholds.9 Therefore myogenic potentials to bone conducted sound were measured in 10 older subjects aged 63 to 83 years (mean (SD), 71.6 (6.7)), using 7 ms/500 and 1000 Hz tones of 10 V p–p (equivalent to 112 and 107.5 dB SPL). These frequencies, at which the bone conductor had its greatest output, were chosen to enable more effective stimulation. Those who had no response to stimuli with these parameters were studied using more intense stimuli of 20 V p–p (115 and 112 dB at 500 and 1000 Hz). Stimulus thresholds were also measured. VEMP to air conducted tones with similar parameters were recorded and thresholds measured for comparison.
We measured responses (net potentials) to combined air and bone conducted tones delivered to the same ear at varying degrees of phase advance of the bone conducted tone, using 20° increments. Air and bone conducted tones (7 ms/500 Hz) with intensities that evoked responses of comparable amplitudes were used. The amplitude of the net potential to the combined stimuli was measured. Decreases in the net potential were interpreted as evidence of phase cancellation of the stimuli.
Patients with specific lesions
Responses to clicks (145 dB SPL) and to air and bone conducted tones with the standard parameters were measured in four subjects who had undergone selective unilateral vestibular neurectomy, in six subjects with conductive hearing loss (“air–bone gaps” of 10–40 dB), and in four subjects with profound sensorineural hearing loss. Four subjects (five ears) with the “Tullio” phenomenon secondary to superior semicircular canal dehiscence had stimulus thresholds measured to clicks, air conducted tones, and bone conducted tones. One subject with a bilateral near total vestibulopathy, who had marked reduction of caloric responses and decreased VOR in the planes of all six semicircular canals (Cremer P, personal communication), was studied using air conducted clicks and tones and bone conducted tones.
VEMP evoked by air conducted clicks, tones, and forehead taps
VEMP to clicks and tone bursts delivered through headphones were measured in all 10 subjects. The clicks of 0.1 ms duration and 145 dB SPL (= 100 dB NHL) were generated using an ST 10 evoked potential stimulator (Medelec, Surrey, UK) and tone bursts of 2 V p–p intensity (= 123, 123.5, 124, and 125 dB SPL at 250, 500, 1000, and 2000 Hz) were generated using the function generator. During each recording, stimuli were presented pseudorandomly in each ear at five per second through calibrated headphones (TDH 49, Telephonics Corp, New York, USA) and responses to 512 stimuli (256 in each ear) were averaged. Frequency tuning properties of air conducted tones were examined using the same methods as for bone conducted tones. Click and air conducted tone thresholds were measured using 5 dB and 6 dB decrements, respectively.
Forehead taps were delivered manually through a small gauze pad placed approximately over Fpz, using a nylon handled reflex hammer fitted with an inertial trigger switch. Responses to 64 stimuli were averaged. In subjects who did not have two separate negative peaks (n1,n2), a slur on the upstroke leading to the negativity or a point at which a clear change in the slope of the upstroke occurred was marked as n1. Data of subjects who had “non-separating peaks” without these features were treated as missing values.
Statistical methods
SPSS version 9 (SPSS Inc, Chicago, Illinois, USA) was used for all data analyses. To examine the effects of varying location and frequency, repeated measures analysis of variance (ANOVA) with contrasts were done using the corrected reflex amplitudes as the dependent variables and stimulus location or stimulus frequency as fixed factors. A significance level of 0.05 was used, with Bonferroni corrections for multiple comparisons. All results are given as mean (SD) unless otherwise noted.
RESULTS
Properties of bone conducted responses and comparison with VEMP to other stimuli
All 10 subjects had short latency biphasic positive–negative (p1n1) responses to bone conducted tone bursts of standard parameters, which were present bilaterally. Later responses were also present but will not be discussed further. Eight subjects had larger amplitude bone conducted responses recorded from the SCM ipsilateral to the stimulus and smaller ones from the contralateral SCM (see fig 1) and two had the reverse. All subjects had VEMP to clicks and “air conducted” tone bursts of standard parameters recorded from the SCM ipsilateral to the side of stimulation. Seven of the 10 subjects had measurable VEMP to forehead taps recorded from both SCM, two had measurable VEMP on one side (SCM) only. There were four “non-separating” responses (two unilateral and one bilateral).
Bone conducted tones evoked myogenic responses, which had significantly higher average corrected amplitudes (mean (SD), 1.36 (0.45)) recorded from the SCM ipsilateral to the stimulus compared with the contralateral SCM (0.93 (0.66)) (p < 0.001). Raw amplitudes were 77.0 (26.2) μV ipsilaterally and 55.2 (34.8) μV contralaterally. Peak latencies recorded from the SCM ipsilateral to the stimulus occurred earlier (average p1 and n1 of 13.6 (1.8) ms and 22.3 (1.2) ms) compared with those recorded over the contralateral SCM (p1 and n1 of 14.9 (2.1) and 23.7 (2.7) ms). The range for p1 latencies was 11.9 to 16.5 ms ipsilaterally and 12.5 to 19.6 ms contralaterally, and that for n1 latencies was 20.0 to 26.6 ms ipsilaterally and 20.1 to 30.2 ms contralaterally.
The average corrected amplitudes for click, tap, and air conducted tone burst evoked responses were 1.98 (0.48), 2.4 (0.52), and 1.3 (0.52) μV (raw amplitudes of 130.4 (32.9), 247.0 (61.2), and 78.3 (32.8) μV, respectively). The average peak latencies (p13n23 or p1n1) for clicks, taps, and air conducted tones were similar to those reported in previous studies (12.4 (1.2) ms and 20.6 (1.3) ms for clicks; 9.7 (1.2) ms and 18.2 (1.6) ms for taps; and 14.3 (1.3) and 22.9 (1.7) ms for 500 Hz air conducted tones).5,9
Three of the four subjects who had undergone selective vestibular neurectomy as a treatment for intractable vertigo had absent short latency bone conducted responses in the ipsilateral SCM on stimulation of the mastoid on the side of the lesion. On stimulation of the intact side, p1n1 responses were present on that same side only. One of these three subjects, when stimulated over the operated side, had no response ipsilaterally but a well formed p1n1 response was recorded from the contralateral SCM, presumably due to bone conduction of the tone to the opposite labyrinth (fig 2). A fourth subject who had undergone selective vestibular neurectomy had a small p1n1 response recorded from the ipsilateral SCM following stimulation of the side of the lesion, but this finding was not reproducible on repeated recordings. The subject was also found to have residual posterior semicircular canal function on impulsive testing,1 suggesting residual nerve function related to the inferior division of the vestibular nerve (Cremer P, personal communication). As with click and tone burst evoked VEMP, short latency responses to bone conducted sound were preserved in the four subjects with severe sensorineural hearing loss. The above findings confirm the conclusions of Sheykholeslami et al,8 that short latency p1n1 myogenic responses to bone conducted sound are vestibular dependent and we shall henceforth refer to them as “bone conducted VEMP”.

Myogenic potentials to bone conducted tones after right vestibular neurectomy. Upon stimulating over the right mastoid, no p1n1 response is seen over the right sternocleidomastoid (SCM), although a well formed response is present over the left SCM probably secondary to conduction of the tone across the skull (see text). When the unaffected (left) side is stimulated, a response is observed ipsilaterally but no response is observed on the right. Note that later responses were not abolished on the right.
All five subjects tested for the effect of stimulus location had bone conducted VEMP to the standard stimulus parameters when the bone conductor was placed on the mastoid process (3 cm posterior to the external acoustic meatus). When stimulated over the temporal and occipital regions, ipsilateral bone conducted VEMP were obtainable only in three and two of the five subjects, respectively. Stimulation over Fpz resulted in bilateral responses in three of the five subjects, a unilateral response in one, and absent responses in the other. The average corrected VEMP amplitudes for the mastoid, frontal, anterior temporal, and occipital regions were 1.22, 0.32, 0.36, and 0.31, respectively. Raw amplitudes were 76.0, 44.0, 14.0, and 6.5 μV, respectively. Upon detailed mapping of amplitudes, when the stimulus was applied at 12 different locations in relation to the external acoustic meatus, larger bone conducted VEMP were recorded from locations superior and posterior to the meatus (fig 3). A location 3 cm behind and 2 cm above the external acoustic meatus yielded the highest average VEMP amplitude for the 10 subjects, although the best site varied between subjects. A significant effect of location was present on ANOVA (F11, 99 = 16.5, p < 0.001). On simple contrasts between VEMP amplitudes at the optimum location and the remaining sites, there were significant differences between it and eight of the remaining 11 locations. There were no significant differences between the amplitude at the optimum site and those at three other locations (“optimum locations”). These formed an inverted L behind and above the external acoustic meatus and were located 3 cm behind, 3 cm behind and 2 cm above, 3 cm behind and 4 cm above, and 4 cm directly above the external acoustic meatus, respectively (fig 3).

The effect of stimulus location on bone conducted vestibular evoked myogenic potential (VEMP) amplitude. The upper half of the figure shows individual VEMP recorded from one subject’s left sternocleidomastoid (SCM) with the bone conductor placed on the left mastoid process and the adjacent temporal bone over 12 locations adjacent to the external acoustic meatus. The bar graph gives average VEMP amplitudes for 10 subjects at each of these locations. The four highest mean amplitudes (pale bars) were observed at the four locations (each marked with an asterisk) closest to the external acoustic meatus posteriorly and superiorly.
The peak to peak amplitudes of bone conducted VEMP increased linearly with increasing mean rectified EMG activity (fig 4). Correlation coefficients (r) between SCM activation and peak to peak amplitudes in the same subject ranged from 0.9 to 0.99. In the subject who was tested at 15 different levels of background activation (18 to 155 μV), the raw peak to peak amplitudes varied from 21.1 to 262 μV (12-fold). The corrected amplitudes only varied from 1.1 to 1.9 (1.7-fold), confirming that this measure was more constant and less likely to be influenced by EMG activity when compared with raw amplitudes. Correlation of VEMP amplitudes to the standard stimulus obtained on two separate days (approximately one week apart) for 10 subjects resulted in a coefficient of 0.77 (p < 0.001) for bone conducted VEMP (n = 10). A similar correlation coefficient (0.71) was obtained for tap evoked responses (n = 10). The tendency to have larger VEMP recorded from the contralateral SCM, observed in two subjects, was not reproducible between experiments.

The effect of background electromyographic (EMG) activity on bone conducted vestibular evoked myogenic potential (VEMP) amplitudes. The traces are averaged unrectified EMG recorded from the right sternocleidomastoid (SCM) of one subject during different degrees of activation in response to a 112 dB SPL (10 V p–p)/500 Hz/7 ms bone conducted tone delivered to the right mastoid process. The mean rectified EMG activity level for the 20 ms preceding the stimulus is given on the left side, next to the raw traces. Peak to peak amplitudes rise linearly with background activation (r2=0.92).
Frequency tuning properties and side to side differences
Average corrected VEMP amplitudes recorded from the ipsilateral SCM for 7 ms/250, 500, 1000, and 2000 Hz bone conducted tones (107 dB nominally) were 1.43 (0.2), 0.68 (0.2), 0.88 (0.2), and 0.30 (0.1), respectively (fig 5). Average raw amplitudes were 104.4 (60.2), 58.1 (54.7), 70.2 (47.0), and 24.1 (15.9) μV, respectively. Differences between reflex amplitudes measured at the four stimulus frequencies were much greater than the slight differences in stimulus intensity for each frequency (see Methods). In contrast to air conducted tones (fig 5), the largest response was at 250 Hz. On ANOVA with contrasts, average VEMP amplitudes in response to the 250 Hz stimulus were significantly higher than average amplitudes obtained with the other three stimuli (F3,27 = 41.64, p < 0.001). There was no significant difference between responses to 500 and 1000 Hz stimuli (p = 0.56). The optimum stimulus frequency was 250 Hz for 17 sides and 1000 Hz for three sides. Mean thresholds for bone conducted tones were 97.5 (3.9) dB SPL (30.5 dB HL (hearing level)) when measured using the 250 Hz stimulus and 100.3 (5.0) dB SPL (42.3 dB HL) using a 500 Hz stimulus. Thresholds for clicks and air conducted tones were significantly higher at 131.7 (4.9) dB SPL (86.7 dB NHL) and 114 (5.3) dB SPL (106 dB HL), respectively (fig 6).
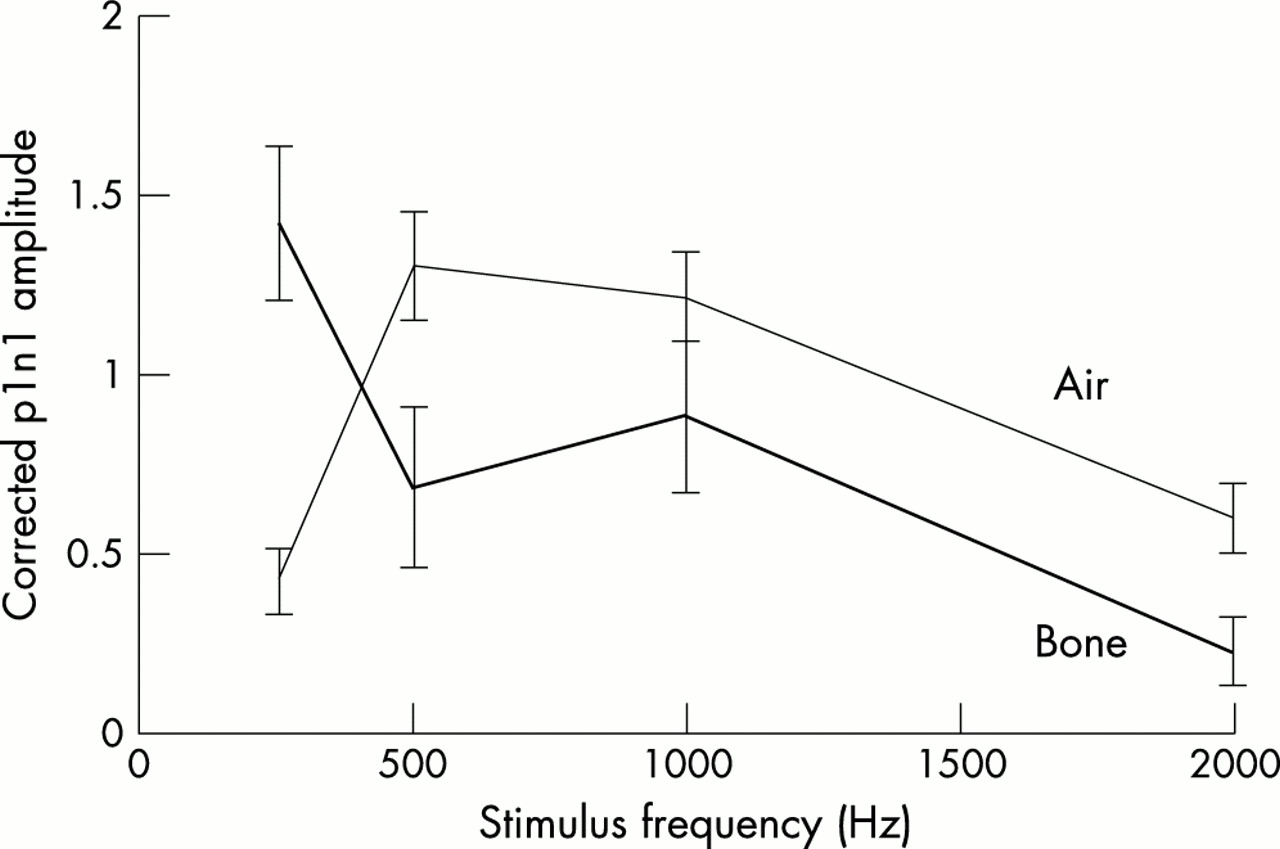
Frequency tuning properties of air and bone conducted tone burst evoked responses. Mean and SEM values of vestibular evoked myogenic potential (VEMP) amplitudes in response to 7 ms tone bursts of 250, 500, 1000, and 2000 Hz are given; the lines connect average amplitudes at the four frequencies. The thin line represents VEMP to air conducted tone bursts of 123 to 125 dB SPL and the thick line, VEMP to 105.5 to 107.5 dB bone conducted tone bursts. Air conducted tones have a tuning curve that peaks between 500 and 1000 Hz with marked reduction of mean VEMP amplitudes at 250 Hz. Bone conducted VEMP, in contrast, have maximal amplitudes at 250 Hz.

Thresholds to three modes of stimulation. Normal subjects had click thresholds ranging from 120 to 135 dB SPL, air conducted tone thresholds of 106 to 124 dB SPL, and bone conducted thresholds of 95 to 107.5 dB SPL at 500 Hz. The click thresholds for subjects with the Tullio phenomenon were 105 to 110 dB SPL and air and bone conducted thresholds were 88 to 94 dB SPL and 83 to 95 dB SPL, respectively. While the click and air conducted tone thresholds in Tullio subjects were at least 10 dB below the lowest normal threshold, those for bone conducted tones approached normal levels in two of the five ears tested.
Asymmetry ratios for bone conducted tones were calculated using the VEMP amplitudes recorded from the SCM ipsilateral to the stimulus. The average asymmetry ratios for bone conducted tones were 17.3% (range 0 to 58.2), 38.5% (0 to 100), 47.8% (0 to 100), and 76.9% (0 to 100) at 250, 500, 1000, and 2000 Hz, respectively. Those for air conducted tones were 43.1% (0 to 100), 24.9% (0 to 62.7), 14.6% (0 to 35.5), and 42.8% (0 to 100) at 250, 500, 1000, and 2000 Hz. One subject with a short thick neck and SCM muscles that were difficult to palpate had absent bone conducted VEMP to all but the 250 Hz stimulus (at an intensity of 107 dB), and this was responsible for the wide range of asymmetry ratios obtained for the 500 to 2000 Hz frequencies. The mean asymmetry ratios for VEMP to clicks and taps were 5.4% (0 to 14.4) and 21.1% (0 to 42.8).
In the 10 older subjects tested (20 ears), six sides had absent VEMP to 500 Hz and 1000 Hz bone conducted stimuli of 10 V peak to peak (equivalent to 112 and 107.5 dB SPL, respectively). All these had detectable VEMP when the stimulus was increased to 20V peak to peak (115 and 112 dB at 500 Hz and 1 kHz, respectively). VEMP to air conducted tones were absent in seven and five ears at frequencies of 500 and 1000 Hz, respectively. None of these subjects had conductive hearing loss on audiometric testing. Stimulus thresholds at 500 Hz were 108.6 (4.6) dB SPL (50.6 dB HL) for bone conducted VEMP (n = 20 ears) and 121.5 (2.9) dB SPL (113.5 dB HL) for air conducted tones (n = 15 ears), both of which were significantly higher than the thresholds obtained in the 10 younger subjects using 500 Hz stimuli (p < 0.001 in both cases).
We were able to demonstrate modest phase cancellation between air and bone conducted tone burst evoked responses. For the three subjects tested, average reflex amplitudes to the air conducted stimulus alone, bone conducted sound alone, and the maximum net potential obtained with the combined stimulus were 1.01, 1.06, and 2.38, respectively. Thus the maximum net potential approximated the sum of the responses to air and bone conducted sound. For subject 1, the maximum corrected amplitude for the net potential was 2.77 at intervals of 60° and 120°; the minimum amplitude was 1.8 at 300° (35% attenuation). For subject 2, the corresponding values were 1.77 at 80° and 1.26 at 280° (29% attenuation), and for subject 3, 2.6 at intervals of 60° and 160° and 1.28 at 320° (51% attenuation). The amplitudes of the net potentials were normalised to the average net potential of each subject and plotted against the phase interval between the two stimuli (fig 7). A tendency for the net potential to decrease between phase differences of 90° and 270° was observed.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Phase cancellation of air conducted sound evoked responses by bone conducted sound. The scatter plot demonstrates the amplitudes of the “net potentials” of three subjects stimulated by air and bone conducted tones delivered in the right ear at varying phase intervals between 20° and 360° (the phase of the tone delivered by the bone conductor always preceded that delivered by the headphones). All amplitudes have been normalised to the individual’s average reflex amplitude (average net potential). The line, which represents a LOWESS fit of the data, shows a fall in the amplitude of the net potential between phase differences of 90° and 270°.
Subjects with specific lesions
Six subjects with conductive hearing loss (eight affected ears) had absent VEMP to 145 dB SPL clicks and air conducted tone bursts of standard parameters on the affected sides. In contrast, VEMP to bone conducted tones of 115 dB/500 Hz/7 ms were present for in all affected ears.
Four subjects (five ears) with the Tullio phenomenon secondary to dehiscence of the superior semicircular canal10 had average thresholds of 107 dB SPL (62 dB NHL) to clicks, 90.4 dB SPL (82.4 dB HL), to air conducted tones, and 90.2 dB SPL (32.2 dB HL) to bone conducted tones, all significantly lower than those of normal subjects (p < 0.001). The average reduction in thresholds from normal values was 24.7, 23.6, and 10.1 dB for air conducted clicks and tones and bone conducted tones, respectively (fig 6).
The subject with markedly reduced caloric responses binaurally (maximum SPV on stimulation of either side = 2°/s) as well as marked reduction of VOR to head impulses in the planes of all three semicircular canals on each side, had preservation of VEMP to clicks, air conducted tone bursts, and bone conducted tone bursts of standard parameters delivered to the right side, with corrected amplitudes of 0.97, 1.43, and 0.76 and raw amplitudes of 52.5, 44.9, and 45.1 μV, respectively. Upon stimulation of the left ear, VEMP to clicks were absent, and responses to air and bone conducted tones were markedly reduced, with corrected amplitudes of 0.37 and 0.42 and raw amplitudes of 23.8 and 67.0 μV, respectively.
DISCUSSION
Although head rotation and acceleration are the adequate physiological stimuli for the vestibular apparatus, non-mechanical stimuli such as loud sound, vibration, and galvanic currents activate primary vestibular afferents and have been used to evoke VEMP.3,7,11 Vestibular responses to sound have been attributed to stimulation of the saccule, which in other species such as fish and amphibians serves as a primary or secondary organ of hearing.12,13 Recordings from primary and secondary vestibular neurones in guinea pigs and cats in response to intense clicks or tones have shown increases in neural discharge.14–16 Using labelling techniques and responses to physiological stimuli, acoustically responsive afferents been traced to the saccule.15 The vestibular apparatus is sensitive to vibration in amphibians17,18 and mammals.19–22 In squirrel monkeys, both audio frequency sound and vibration can activate primary afferents from all vestibular end organs, with saccular units showing greater sensitivity than those from the canals to acoustic stimuli but not to vibration.19 Vestibular responses such as illusions of self motion and nystagmus following direct vibration of the head using 120–280 Hz stimuli were reported by Lackner and Graybiel,20 who attributed these effects to stimulation of the semicircular canals. More recently, low frequency (100 Hz) vibration of the mastoid process has been found to evoke nystagmus (beating away from the side of the lesion) and shift of the subjective visual horizontal (towards the lesion) in subjects with unilateral vestibular hypofunction,21,22 which may reflect asymmetrical vestibular activation.
Bone conducted sound of 7 ms/500 Hz/112 dB SPL produced short latency p1n1 potentials in all 10 normal subjects aged 24 to 52 years, responses which were abolished following selective vestibular neurectomy, and preserved despite profound sensorineural hearing loss, and therefore appear to be another form of VEMP. Later responses, sometimes present, were not abolished by vestibular nerve section. Unlike VEMP to clicks and air conducted tones, bone conducted VEMP were often bilateral. Most subjects had larger bone conducted VEMP with earlier peak latencies (p1 and n1 ranging between 11.9 and 16.5 ms and between 20.0 and 26.6 ms) recorded from the ipsilateral SCM and smaller ones with average peak latencies occurring about 1 ms later contralaterally (p1 and n1 ranging between 12.5 and 19.6 ms and between 20.1 and 30.2 ms). Bone vibration spreads widely throughout the skull, and the contralateral bone conducted VEMP are likely to be responses arising from the opposite labyrinth following stimulus conduction across the skull. Although crossed neural pathways are known to exist,11 contralateral bone conducted VEMP clearly differ from the “n1p1” crossed neural responses to galvanic stimulation,11 which are of opposite polarity (negative–positive responses) and also have earlier peak latencies compared with the ipsilateral response. The findings in the subject with selective vestibular nerve section indicate that “crossed” bone conducted VEMP do not depend upon activation of the ipsilateral vestibular apparatus. The larger bone conducted VEMP recorded from the contralateral SCM in two subjects further illustrate the vagaries of sound transmission by bone conduction. Similar observations have been made for hearing. Bone conducted sound delivered through the mastoid process has been shown, owing to the asymmetry of skull thickness, to stimulate the contralateral cochlea more effectively in some subjects.23 The optimum stimulus frequency for bone conducted VEMP was 250 Hz for most of the ears tested. Similar optimum frequencies have been reported by Sheykholeslami et al.24 Differences between air and bone conducted tone bursts are likely to relate to the different filtering properties of bone and the middle ear as well as to potential differences in the mechanism and specific site of action of the two stimuli. As the B71 vibrator did not permit delivery and accurate calibration of tones of less than 250 Hz frequency, we were unable to define the tuning curve fully.
While VEMP to air conducted sound are thought to be of saccular origin, the precise origin of the afferents responsible for the bone conducted VEMP is not yet known. Previous experiments which achieved phase cancellation of an air conducted sound by a bone conducted sound25,26 demonstrated that the two stimuli activate cochlear receptors by similar mechanisms. We have shown that the bone conducted VEMP is vestibular dependent and has similar latency and waveform to the VEMP evoked by air tone bursts. Results obtained from the patient with markedly attenuated canal function imply that bone conducted VEMP arise from the otolith organs. Partial cancellation of responses when air and bone conducted tones were delivered out of phase is consistent with overlap in the receptors stimulated by the two methods. We conclude that the receptors responsible for bone conducted responses arise from the otoliths and in part, at least, from the saccule.
Stimulation over the mastoid yielded bone conducted VEMP more consistently compared with frontal and occipital bones and the temporal bone anterior to the external acoustic meatus. Audiometric studies comparing bone conduction thresholds over the frontal bones and mastoid report lower thresholds over the mastoid.27–30 Studies in humans comparing BAER latencies with the stimulus over the frontal, occipital, and temporal bones have reported earlier wave V latencies to temporal stimulation.31,32 Our optimum location for stimulus delivery occurred almost always in one of the four locations forming an inverted “L” shape posterior and superior to the external acoustic meatus; other locations near the meatus gave smaller responses. Sohmer and Freeman reported shorter BAER latencies when the bone vibrator was placed anterior to the external acoustic meatus over the thinnest part of the temporal bone.32 We did not find bone conducted VEMP amplitudes to be significantly larger anterior to the external acoustic meatus. This may reflect the more posterior location of the vestibular apparatus compared with the cochlea. Our results indicate that effective stimulation can be delivered at any one of the four optimal locations providing the bone conductor makes good mechanical contact. Symmetrical sites should be used to minimise differences in amplitude between sides.
The perceived loudness of sounds is a function of the middle ear, the cochlea, and the central neural pathways. The standard measure of this property, for continuous sound, is expressed as dB HL (hearing level) and the relation between dB HL and dB SPL (sound pressure level) differs with frequency. The HL reference is not appropriate when the cochlea is not the organ being stimulated by sound, as is the case for VEMP. The reference levels for air and bone conducted sound (0 dB) are defined differently. For sound in air, 0 dB SPL is defined as a pressure (20 μP), for bone conduction 0 dB SPL is defined as a force (1 μN). The different reference levels prevents direct comparison of air and bone conducted sound. If force is used for comparison, 0 dB for air is substantially less than 1 μN. At 1 kHz, 0 dB HL (for sound in air) is close to 0 dB SPL, implying an extraordinary level of sensitivity, close to the physical limits of detection. Contrasting with sound in air, 0 dB HL for continuous bone conducted sound is 42.5 dB higher than the SPL reference for 1 kHz (and therefore approximately 130 times less sensitive). von Békésy33 argued that this reduced sensitivity is a desirable characteristic that is in part a consequence of the structure of the cochlea.
The stimulus threshold for air conducted VEMP was 114 dB SPL = 106 dB HL, and that for bone conducted VEMP, 97.5 dB SPL = 30.5 dB HL at 250 Hz (using standard HL thresholds).34 The difference between hearing thresholds and VEMP thresholds is much larger for air conducted sound than for bone vibration. Bone vibration at a given perceptual intensity is therefore a more effective vestibular stimulus than air conducted sound, implying a relatively greater sensitivity of the vestibular apparatus to bone conducted sound when compared with the cochlea. Subjects with sound induced vestibular symptoms—the Tullio phenomenon—have a low stimulus threshold for VEMP to air conducted clicks, and this has been attributed to an increased sensitivity of the vestibular apparatus to sound at the receptor level, as neural excitability has been shown to be normal in these subjects.35 Bone conducted VEMP thresholds, although still significantly lower than those of normal subjects, were not as markedly reduced as the thresholds of VEMP to air conducted clicks in the six subjects with the Tullio phenomenon. A significant proportion of “bone conduction” does in fact travel through the middle ear26,36 so that some reduction in bone conducted VEMP thresholds would therefore be expected. The vestibular receptors themselves appear to be normal; thus the component of vestibular activation caused by a direct effect on the receptors by distortional vibration and inertial effects on the otoliths may be unaltered, leading to lesser abnormalities than occur with air conducted sound.
The clinical indications for the use of bone conducted VEMP are similar to those for taps, and they have a similar reproducibility, but they are more consistently present in normal subjects and can be calibrated. Bone conducted VEMP are therefore a suitable alternative to the tap VEMP but they do not replace click evoked VEMP, which are more symmetrical, are highly reproducible (r = 0.9) between experiments,9 and have a wider separation between normal thresholds and those of subjects with the Tullio phenomenon. All subjects with absent click evoked responses should be tested for VEMP to either bone conducted tones or taps to exclude conductive hearing loss as the cause. When compared with air conducted sound, larger bone conducted stimuli may be safely used, as they do not stimulate the cochlea as effectively. This technique may also be useful in studying older subjects with higher stimulus thresholds.9 Absent VEMP to both air and bone conducted sound suggest a lesion along the vestibulocollic reflex pathway. Measurement of VEMP to transmastoid galvanic stimulation11 may yield useful information in this context, as this stimulus acts upon the terminal part of the primary vestibular afferents. The preservation of a response to galvanic stimulation when sound evoked responses are absent makes a lesion of the vestibular end organ most likely.37
Conclusions
Bone conducted sound evokes VEMP that provide an additional measure of vestibular function, which is not confounded by conductive hearing loss. They complement information obtained from click VEMP and other techniques currently used in vestibular assessment.
Acknowledgments
This research was supported by a grant from the National Health and Medical Research Council of Australia. MSW received a medical research fellowship from the Garnett Passe and Rodney Williams Memorial Foundation during the period of this study. SMR translated excerpts of the work of Tullio. We thank Dr Toshihisa Murofushi for introducing the technique of measuring bone conducted VEMP, Ms C Urban for translating the work of von Békésy, Dr G Colin-Thome for audiological advice, Mr P Bosloper for calibrating the bone conductor, and Mr Chi Sum Tsang for assistance with patient recruitment. We also acknowledge the expert assistance of Drs Phillip Cremer and Americo Migliaccio in testing three dimensional VOR of two subjects with specific lesions.
REFERENCES
Footnotes
-
Competing interests: none declared
Linked Articles
- Correction