Article Text

Abstract
Background and objective Aetiology and pathogenesis of anti-N-methyl-D-aspartate receptor (anti-NMDAR) encephalitis, the most common autoimmune encephalitis, is largely unknown. Since an association of the disease with the human leucocyte antigen (HLA) has not been shown so far, we here investigated whether anti-NMDAR encephalitis is associated with the HLA locus.
Methods HLA loci of 61 patients with anti-NMDAR encephalitis and 571 healthy controls from the Chinese Han population were genotyped and analysed for this study.
Results Our results show that the DRB1*16:02 allele is associated with anti-NMDAR encephalitis (OR 3.416, 95% CI 1.817 to 6.174, p=8.9×10−5, padj=0.021), with a higher allele frequency in patients (14.75%) than in controls (4.82%). This association was found to be independent of tumour formation. Besides disease susceptibility, DRB1*16:02 is also related to the clinical outcome of patients during treatment, where patients with DRB1*16:02 showed a lower therapeutic response to the treatment than patients with other HLA alleles (p=0.033). Bioinformatic analysis using HLA peptide-binding prediction algorithms and computational docking suggested a close relationship between the NR1 subunit of NMDAR and the DRB1*16:02.
Conclusions This study for the first time demonstrates an association between specific HLA class II alleles and anti-NMDAR encephalitis, providing novel insights into the pathomechanism of the disease.
Statistics from Altmetric.com
Introduction
Autoimmune encephalitis (AE) refers to a newly described group of diseases caused by autoantibodies against neuronal protein. Clinically, it is a severe and rare autoimmune disorder characterised by psychiatric symptoms, seizures, encephalopathy, cognitive and movement impairment.1 Among the group of AE, the most common member is the anti-N-methyl-D-aspartate receptor (NMDAR) encephalitis which is featured by autoantibodies against the GluN1 (NR1) subunit of NMDAR.2 3 The aetiology of anti-NMDAR encephalitis is still not clear, although both tumour formation1 4 5 and viral infection1 6–8 have been suggested as causative factors of the disease. Anti-NMDAR encephalitis was identified in 20075 and its pathogenesis has been extensively investigated since then. It is believed that binding of autoantibodies to the NR1 subunit of NMDAR mediates receptor internalisation leading to a decrease in its surface density and synaptic localisation of NMDAR clusters which ultimately results in glutamatergic synaptic dysfunction.2 Although the principle of the autoantibody-mediated dysfunction of neuronal cells is relatively clear, mechanisms involved in the induction and control to the anti-NMDAR autoantibodies themselves remain elusive.
Given that AE is associated with autoantibodies against neuronal proteins, it is conceivable that human leucocyte antigen (HLA) class II loci are associated with the susceptibility to AE. In consistence with this hypothesis, anti-leucine-rich glioma-inactivated 1 encephalitis, another common AE, has recently been reported in three independent studies to be associated with the class II allele HLA-DR7.9–11 Unexpectedly, no association to HLA class II could be demonstrated for anti-NMDAR encephalitis.10 11 With a very small number of patient samples (n=17), Kim et al failed to show any significant association between a specific HLA locus and anti-NMDAR encephalitis.10 In a genome-wide association study including 96 patients and 1194 controls, Mueller and colleagues demonstrated that anti-NMDAR encephalitis was associated with several single nucleotide polymorphisms (SNP) located within the HLA I locus, however, no disease-associated SNPs could be identified within the HLA II locus.11
In this study, we aimed to analyse the potential association between HLA locus and anti-NMDAR encephalitis with a large cohort of patients and healthy individuals. We genotyped 61 patients and 571 healthy controls in three HLA class I genes (HLA-A, B and C) and five HLA class II genes (DRB1, DQA1, DQB1, DPA1, DPB1), and correlated the association of HLA locus with disease susceptibility and clinical features of anti-NMDAR encephalitis.
Subjects and methods
Subjects
Sixty-one consecutive patients with anti-NMDAR encephalitis were recruited at the Department of Neurology, the Third Affiliated Hospital of Sun Yat-sen University, Guangzhou, China. All patients were hospitalised at the Department of Neurology between March 2014 and May 2018 and diagnosed based on the criteria according to Graus et al.12 To test autoantibodies against NMDAR, patients’ cerebrospinal fluid (CSF) samples were analysed by cell-based indirect immunostaining using a commercially available kit (EUROIMMUN Medizinische Labordiagnostika, Lübeck, Germany), as described previously.13 No patient had comorbidity with other autoimmune diseases. All patients were screened for systemic tumours with ultrasonography, CT or/and MRI. Outcomes of the neurological status of each patient were assessed using the modified Rankin Scale (mRS) with a score ranging from 0 to 5, characterised by no symptoms (0); no significant disability despite symptoms (1); slight disability (2); moderate disability (3); moderate-to-severe disability (4); and severe disability (5).14 Patients were treated with a first-line drug regimen (steroids, intravenous immunoglobulins) alone or in combination with a second-line regimen (rituximab, azathioprine or cyclophosphamide). Patients were followed up 3 months after treatment and received repeated assessments of mRS scores at the Third Affiliated Hospital of Sun Yat-sen University. Clinical evaluation was performed by an experienced neurologist who was blinded to the treatment and genotyping results of the patients. We reviewed the demographic and clinical information of the patients, the MRI of the brain and the CSF examinations.
DNA preparation and HLA genotyping
Blood samples from all patients were collected and stored at −80°C. Genomic DNA was extracted from peripheral blood leucocytes using a GenMag Blood Genome DNA Extraction Kit according to the manufacturer’s recommendations. The genotypes of the HLA class I (HLA-A, B and C) and class II (DRB1, DQA1, DQB1, DPA1, DPB1) genes of each patient were determined at the four-digit allele level using a PCR sequence-based typing method, as previously described.15 16 HLA genotyping was also performed in samples derived from 571 geographically and ethnically matched healthy controls, including 90 healthy controls from the Medical Examination Centre of the Third Affiliated Hospital of Sun Yat-sen University, Guangzhou, China, and 481 ethnic Hans from southern China as previously reported by us.15
Statistical analyses
To assess the significance of clinical variables between groups, the Student’s t-test was applied for normally distributed data, while Mann-Whitney U test was performed for data lacking normal distribution. Differences in genotype frequencies of HLA alleles or haplotypes were analysed by Fisher’s exact test. The p values determined by Fisher’s exact test were adjusted for multiple testing with the false discovery rate step-up method, only the adjusted p values (padj) <0.05 (two tailed) were considered as statistically significant. The statistical analyses were performed using R-studio software and GraphPad Prism V.5.0. In control groups, the presence of the Hardy-Weinberg equilibrium was determined using the method according to Guo and Thompson.17
HLA peptide-binding prediction and in silico docking
The NetMHCIIpan 3.2 Server, which represents the currently most accurate tool for predicting major histocompatibility complex (MHC) class II peptide-binding affinities, was used to predict epitopes.18 The NR1 protein sequence obtained from the National Center for Biotechnology Information (accession number NP_015566.1) was submitted to the server. DRB1*16:02 was selected for predicting epitopes within the protein. Thresholds for weak and strong binding peptides were set as 1% and 10% ranks, respectively. The in silico docking of the NR1 segment into DRB1*16:02 was performed using the AutoDock Vina software with the docking grid encompassing the entire peptide-binding cleft and the option ‘number of torsions’ set to zero.19 The prediction of the tertiary structure and the docking of the peptide were performed as described previously.10 In brief, AutoDock Tools was used to add hydrogens, to assign charges and to specify rotatable bonds to the proteins.20
Data availability statement
Anonymised data not published within this article will be shared by request from any qualified investigator.
Results
Demographic and clinical status of the patients with anti-NMDAR encephalitis
Sixty-one patients with anti-NMDAR encephalitis (31 females and 30 males) were enrolled in this study. The age of the patients ranged from 4 to 64 years (mean age of 25.6 years). Of the 61 patients with anti-NMDAR encephalitis, 7 (11.5%) showed CSF virus antibody positivity, including herpes simplex virus (HSV)-1 IgG positivity (n=2), and HSV-1/2 IgG positivity (n=4), cytomegalovirus IgG positivity (n=1). Table 1 summarises the clinical features of the patients, including disease onset, mRS scores, MRI abnormalities, tumour status and disease symptoms. All patients received first-line treatment with or without combination with second-line treatment. Tumour removal was performed in 12 out of 15 patients with tumour.
Demographic and clinical status of patients with anti-NMDAR encephalitis
Anti-NMDAR encephalitis is associated with HLA-DRB1*16:02
In a first step, we compared the frequencies of HLA class I and II alleles between 61 patients and 571 healthy controls. Among all HLA class I and II alleles, a significant association could only be identified between DRB1*16:02 and anti-NMDAR encephalitis. The allele frequency of DRB1*16:02 in patients was 14.75% (18/122), while that in controls (55/1142) was only 4.82% (OR 3.416, 95% CI 1.817 to 6.174, p=8.9×10−5, padj= 0.021, table 2, online supplementary table 1). When a comparison between 46 patients with anti-NMDAR encephalitis without tumour and 571 healthy controls was performed, a significant association was identified between disease and DRB1*16:02 (OR 3.541, 95% CI 1.739 to 6.812, p=0.000313, padj=0.0368) as well as DQA1*01:02 (OR 2.361, 95% CI 1.461 to 3.768, p=0.000284, padj=0.0368) (online supplementary table 2).
Supplemental material
Analysis of HLA class I and II alleles in anti-NMDAR encephalitis and healthy control
Since the two disease-associated HLA class II alleles DRB1*16:02 and DQA1*01:02 form a haplotype,21 we further investigated the association by determining the relationship between four combinations of DRB1*16:02 and DQA1*01:02 with the disease. As shown in table 3, the DRB1*16: 02 + DQA1*01:02 + , but not DRB1*16:02− DQA1*01:02 + , was associated with anti-NMDAR encephalitis, suggesting that indeed DRB1*16:02, but not DQA1*01:02, is associated with the disease.
Association of the DRB1*16:02-DQA1*01:02 haplotype with anti-NMDAR encephalitis
The association of DRB1*16:02 with anti-NMDAR encephalitis is not dependent on the manifestation of tumours
Since some patients with encephalitis developed tumours,1 4 5 it can be assumed that pathogenesis could differ between tumour-positive and tumour-negative patients. Therefore, we next examined whether the association of DRB1*16:02 with anti-NMDAR encephalitis is dependent on the tumour status. Notably, carrier frequencies of DRB1*16:02 in tumour (4/15, 26.67%) and tumour-free patients (13/46, 28.26%) were comparable. Nonetheless, in both groups the frequency surmounted significantly that observed in healthy controls (54/571, 9.46%) (figure 1A). Similar results were obtained if the allele frequency of DRB1*16:02 was compared between patients with or without tumours and healthy controls (figure 1B). Our findings indicate that the association between DRB1*16:02 and anti-NMDAR encephalitis is not affected by the tumour status in the patients.
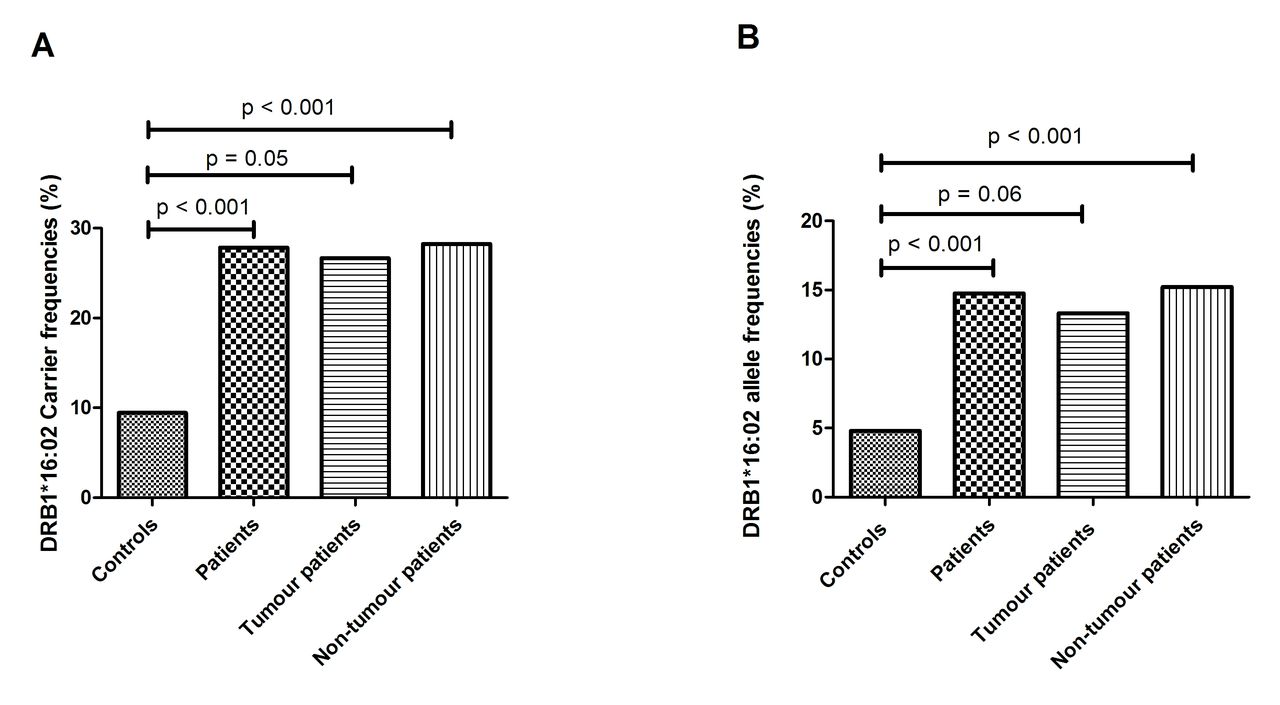
Distribution of the DRB1*16:02 allele in controls and patients with anti-N-methyl-D-aspartate receptor (anti-NMDAR) encephalitis. Carrier (A) and allele (B) frequencies of the DRB1*16:02 in controls, all patients with anti-NMDAR encephalitis, patients with anti-NMDAR encephalitis with tumour and patients with anti-NMDAR encephalitis without tumour. P values were calculated using Fisher’s exact test.
Association of DRB1*16:02 with clinical status of the anti-NMDAR encephalitis
We next investigated whether DRB1*16:02 is associated with clinical features of the anti-NMDAR encephalitis by comparing DRB1*16:02 carriers with DRB1*16:02 non-carriers. As shown in table 4, the two patient subgroups were comparable in age, gender ratio, mRS before the treatment and CSF routine parameters. However, the two subgroups differed significantly from each other in their responses to the treatment determined as reduction of the individual mRS scores. Although both subgroups principally respond to the therapy, the therapeutic effect was significantly lower in patients carrying the DRB1*16:02 allele as compared with the non-carrier patient group. After 3 months of treatment, the median of mRS decrease in patients with DRB1*16:02 reached 1, with a range from 0 to 3, which was found to be significantly lower than that in patients lacking the critical allele (median of decrease of 2, with a range from 0 to 5, respectively; p=0.03, table 4).
Association of the DRB1*16:02 with clinical features of patients with anti-NMDAR encephalitis
Computational epitope prediction and binding of NMDAR to HLA
Since it has been reported that the pathogenic autoantibodies against NMDAR bind to the NR1 subunit,2 3 we tried to predict potential epitopes within this receptor subunit which can be recognised by the DRB1*16:02 protein. By using the NetMHCpan 3.2 Server as a tool for the analysis, we have identified four peptides showing strong binding levels to DRB1*16:02, including ‘FYRIPVLGL’, ‘FLRTVPPYS’, ‘YTANLAAFL’ and ‘FRAITSTLA’ (online supplementary table 3). Among those four predicted epitopes, ‘FRAITSTLA’ showed the strongest binding level to DRB1*16:02. Therefore, we performed further bioinformatics analysis with the predicted epitope of ‘FRAITSTLA’. The position of this epitope with the target sequence ‘FRAITSTLA’ is located between aa880 and aa888 from the N-terminus of NR1 (figure 2A).

{kind=link}
{kind=link}
Bioinformatic analysis of the relationship of the DRB1*1602 and NR1 subunit of NMDAR. (A) Binding affinity prediction of NR1 segment (NMDAR GluN1) to the major histocompatibility complex (MHC) II molecules DRB1*1602. The NetMHCIIpan 3.2 Server was used to predict epitopes within NR1 protein (NP_015566.1). The y-axis presents the binding affinity represented as 1 divided by %Rank of predicted affinity compared with a set of 200 000 random natural peptides, while the x-axis represents the position of amino acid on the NR1 protein. (B) Computational docking of NR1 segment FRAITSTLA to HLA-DRB1*1602 heterodimer. Mesh in the HLA surface indicates close contact between atoms with the NR1 protein segment. HLA, human leucocyte antigen; NMDAR, N-methyl-D-aspartate receptor.
To further evaluate the relationship between the predicted epitope and MHC II molecules, we performed computational docking to describe the binding between the target sequence ‘FRAITSTLA’ and DRB1*16:02. Since the crystallographic structure of DRB1*16:02 has not been experimentally determined, we created its structure by homology modelling with Swiss-Model using the structure of HLA-DRB1*15:01 (Protein Data Bank code 1b×2) as a template.22 As shown in figure 2B, the target sequence ‘FRAITSTLA’ fitted exactly in the peptide-binding groove between HLA-DRB1*16:02 and HLA-DRA1*01:01, with a docking score (∆G) of −6.8 kcal/mol. This in silico docking result suggests a positive interaction between the predicted epitope and DRB1*16:02.
Discussion
In the current study, we have investigated the potential association between specific HLA loci and anti-NMDAR encephalitis. Our results demonstrate for the first time that the HLA class II allele DRB1*16:02 is associated with disease susceptibility and therapeutic response of patients with anti-NMDAR encephalitis. The DRB1*16:02 allele has previously been shown to be associated with an increased risk of multiple autoimmune diseases, including Graves’ disease,23 anti-topoisomerase I autoantibody-positive systemic sclerosis,24 anti-aquaporin 4 antibody-positive neuromyelitis optica spectrum disorder,25 relapsing polychondritis26 and primary Sjögren’s syndrome.27 Interestingly, all of these autoimmune diseases associated with the DRB1*16:02 are predominantly mediated by autoantibodies, suggesting that the DRB1*16:02 as an HLA class II molecule is involved in the autoantibody production.
Notably, our findings on this relation are in contrast to results of two previous studies.10 11 In the study performed by Kim et al in a Korean cohort, the carrier frequencies of the DRB1*16:02 in patients and controls were determined as 5.9% (1/17) and 1.2% (6/485), respectively (OR 5.0, 95% CI 0.6 to 43.9, p=0.225).10 The high OR value in Kim's study is in line with the data of our analysis, although the p value in this study did not reach significance. Besides the very limited number of patient samples (n=17), the extremely low frequency of DRB1*16:02 allele in Korean population could be the main reason for the lack of significant association in Kim's study. The low frequency of the DRB1*16:02 allele in German population28 could also explain the lack of association of DRB1*16:02 with anti-NMDAR encephalitis in Mueller's study.11
Previous reports showed that the presence of a tumour was dependent on age, sex and ethnicity.29 30 The prevalence of tumours in patients with anti-NMDAR encephalitis in the current studies is 24.6%, which is comparable with that in other Chinese populations,31–36 but considerably lower than that in Caucasians.3 5 37 Also, the female-to-male ratios in Chinese patients with anti-NMDAR encephalitis13 31–36 are lower than those in Caucasians.3 5 8 37 This evidence suggests that aetiology of the anti-NMDAR encephalitis might differ slightly between Chinese and Caucasian patients. Interestingly, the current study shows that the association of the DRB1*16:02 allele with the disease does not depend on the presence of tumours. Although different tissues may serve as sources for the autoantigen, our results strongly argue against a source-related effect on the preference of the MHC class II molecules for antigen presentation.
Besides susceptibility, our study also uncovered a relation between the presence of the DRB1*16:02 allele and the therapeutic success of a treatment in patients. Patients expressing the DRB1*16:02 allele recovered to a lower degree after treatment as compared with patients lacking this allele. Association between HLA locus with clinical features of autoimmune diseases has been suggested in some other autoimmune diseases too. For example, the shared epitope, a common structure element of some HLA class II molecules associated with the susceptibility of rheumatoid arthritis (RA), is also a marker of disease severity of RA.38 Another example is that the DRB1*03 allele has been shown to be associated with the clinical response to the treatment of methotrexate in patients with RA.39 Although the precise mechanism behind the association of DRB1*16:02 and patients’ responses to disease treatment remains speculative, two factors might contribute to this phenomenon. On the one hand, disease development in DRB1*16:02 carriers might differ from that in patients lacking this allele. For example, as compared with other HLA II alleles, DRB1*16:02 may recognise a different epitope and thus initiate a different set of autoantibodies against NMDAR, in terms of subclasses and epitope specificity. On the other hand, DRB1*16:02-associated disease development might require a genetic background different from that involved in the absence of that allele, and such genetic difference could contribute to the clinical responses to the treatment.
Interestingly, bioinformatic analysis in the current study predicted a strong epitope for binding to the DRB1*16:02 protein with the target sequence (FRAITSTLA). This target sequence is located within the intracellular C-terminal domain (CTD) of NR1, which plays a critical role in many forms of neurons’ synaptic plasticity,40 41 suggesting that this region contains the major T cell epitope. Although it is known that the major pathogenic B cell epitope is located within an extracellular domain of NR1,2 3 it is conceivable that T cell and B cell epitopes are not located in the same region since antibodies can be generated via linked recognition.42 The role of the epitope with CTD of NR1 in the anti-NMDAR encephalitis needs to be further studied in the future.
Some limitations of this study should be mentioned here. First of all, due to different frequencies of the allele among populations, the association between DRB1*16:02 and anti-NMDAR encephalitis seems to be population specific. Second, since the number of patient samples used in this study is relatively low, these findings need to be confirmed by further studies with a larger number of samples. Third, the treatment of patients enrolled in this study was rather heterogeneous with regard to the therapeutic drugs used and their application schedule. Both parameters are likely to affect the clinical outcome to the treatment. In addition, the mRS scale represents a rather gross estimate for the functional recovery of patients and needs to be explored in the future by more specific psychiatric and neurological outcome measures. Therefore, we plan to evaluate the association of DRB1*16:02 with the clinical response to the treatment in larger groups with more defined treatment and clinical outcomes. Finally, the carrier frequency of DRB1*16:02 allele is rather low (≤30%) in patients with anti-NMDAR encephalitis, suggesting the existence of additional genetic factors predisposing disease which need to be determined in the future.
In conclusion, our study demonstrates that the DRB1*16:02 allele is associated with anti-NMDAR encephalitis, in terms of both disease susceptibility and clinical response to the treatment. These findings suggest that the DRB1*16:02 allele is a risk factor for anti-NMDAR encephalitis and represents a predictive factor for a rather poor clinical response to the treatment. Our results shed some lights on the pathogenesis and treatment strategy of anti-NMDAR encephalitis, especially in populations with high frequencies of the DRB1*16:02 allele.
Acknowledgments
We thank the patients and healthy donors for volunteering to participate in this study.
References
Footnotes
YS and WQ contributed equally.
Contributors Conception and design of the study: YQS, XHY, WQ. Sample collection: YQS, ZHD, CC, XBS, ZQL, YY, FHP, XQH, WQ. Acquisition and analysis of data: YQS, JPY, XYY, JFZ, XLY, SSL. Drafting the text and preparing the figures: YQS, XHY, JFZ, FP.
Funding This study was supported by the National Natural Science Foundation of China (grant numbers: 81771300 and 81701188), the Natural Science Foundation of Guangdong Province, China (grant number: 2017A030313853), the Deutsche Forschungsgemeinschaft, GRK1727 'Modulation of Autoimmunity' and the German Center for Lung Research (DZL).
Competing interests None declared.
Patient consent Obtained.
Ethics approval The Third Affiliated Hospital of Sun Yat-sen University.
Linked Articles
- Editorial commentary