Article Text

Abstract
Objectives: It has been suggested that autoimmunity to peripheral myelin proteins is involved in the pathogenesis of Guillain-Barré syndrome (GBS) and chronic inflammatory demyelinating polyradiculoneuropathy (CIDP). We aimed to compare reactivity of peripheral blood mononuclear cells (PBMC) to antigens of peripheral myelin proteins in patients with GBS and patients with CIDP with that of healthy controls and patients with other non-immune mediated neuropathies (ON).
Methods: We prepared PBMC from blood from 83 healthy controls and from 64 patients with GBS, 54 with CIDP, and 62 with ON. PBMC were tested in antigen specific proliferation assays against peptides from myelin proteins P0, P2, PMP22, and myelin basic protein (MBP), which is identical to myelin P1, and against whole human MBP. Interferon-gamma (IFN-γ) and interleukin (IL)-5 enzyme linked immunospot (ELISPOT) assays were also performed in some subjects to assess spontaneous and peripheral myelin antigen specific PBMC cytokine secretion.
Results: Antigen specific PBMC proliferation assays showed no significant elevation of peptide specific T cell responsiveness in patients with GBS or CIDP compared with healthy controls or patients with ON. Levels of spontaneous ELISPOT IFN-γ secretion were increased in patients with GBS and significantly increased in those with CIDP compared with healthy controls and patients with ON. No convincing differences in antigen specific ELISPOT IFN-γ secretion levels to individual peptides were detectable in patients with GBS. The proportion of patients with CIDP with an increased number of PBMC producing IFN-γ in response to peptide PMP-2251–64 was significantly increased compared with healthy controls and patients with ON. No significant differences in antigen specific ELISPOT IL-5 secretion levels were detectable in patients with GBS or CIDP compared with controls, but levels of spontaneous IL-5 secretion were significantly higher in patients with CIDP than in healthy controls or patients with ON.
Conclusions: Although the lack of significantly increased antigen specific PBMC proliferation in GBS and CIDP does not support a role for T cells, the more sensitive ELISPOT technique detected increased numbers of PBMC secreting IFN-γ spontaneously in 25% of patients with GBS, providing further evidence for a role of T cells in the immunopathology of GBS. Increased numbers of spontaneous IFN-γ and IL-5 secreting cells, and increased IFN-γ secretion in response to PMP-2251–64, in patients with CIDP provide further evidence for a role of myelin specific T cells in CIDP.
- AIDP, acute inflammatory demyelinating polyradiculoneuropathy
- CIDP, chronic inflammatory demyelinating polyradiculoneuropathy
- EAN, experimental autoimmune neuritis
- ELISPOT, enzyme-linked immunospot
- GBS, Guillain-Barré syndrome
- IFN-γ, interferon-gamma
- IL5, interleukin 5
- MBP, myelin basic protein
- ON, other non-immune mediated neuropathies
- PBMC, peripheral blood mononuclear cells
- SFC, spot forming cell
- SI, stimulation index
- TNF, tumour necrosis factor
- Guillain–Barré Syndrome
- Chronic inflammatory demyelinating polyradiculoneuropathy
- ELISPOT
- T-lymphocyte
- autoimmunity
Statistics from Altmetric.com
- AIDP, acute inflammatory demyelinating polyradiculoneuropathy
- CIDP, chronic inflammatory demyelinating polyradiculoneuropathy
- EAN, experimental autoimmune neuritis
- ELISPOT, enzyme-linked immunospot
- GBS, Guillain-Barré syndrome
- IFN-γ, interferon-gamma
- IL5, interleukin 5
- MBP, myelin basic protein
- ON, other non-immune mediated neuropathies
- PBMC, peripheral blood mononuclear cells
- SFC, spot forming cell
- SI, stimulation index
- TNF, tumour necrosis factor
- Guillain–Barré Syndrome
- Chronic inflammatory demyelinating polyradiculoneuropathy
- ELISPOT
- T-lymphocyte
- autoimmunity
The most common form of Guillain-Barré syndrome (GBS) seen in Australia is acute inflammatory demyelinating polyradiculoneuropathy (AIDP), an acute monophasic disease of the peripheral nervous system causing ascending limb weakness. Chronic IDP (CIDP) is a similar disorder, with a chronic relapsing or chronic progressive disease course. Although differing in clinical course, GBS and CIDP have similar pathological findings of primary demyelination and infiltration of peripheral nerves with macrophages and T cells.1,2 The pathogenesis of GBS and CIDP is thought to be due to activation of autoimmune mechanisms by exposure to environmental antigens.3 Infections and immunisations are commonly found to precede episodes of GBS,4,5 and it is thought that these may trigger immune responses that cross react with antigens in peripheral nerves.3,6,7
Immune reactivity to peripheral myelin proteins
Previous studies have attempted to identify peripheral nerve myelin antigen(s) that are the targets of immune attack in GBS and CIDP, including P0, P1 (identical to myelin basic protein (MBP) in the CNS), P2, and PMP-22.8
Some studies have found that patients with GBS and some patients with CIDP have elevated levels of antibody9,10 and T cells specific for P2,11–13 although other studies disagree.14,15 There have been reports of increased antibody to P0 in a minority of patients with GBS,9,10,16 and in one third of patients with CIDP.17 Increased T cell reactivity to P0 in patients with GBS has also been reported.12 Antibody responses to PMP-22 have been described in both inflammatory and non-inflammatory neuropathy,18,19 although this remains controversial.16 There are no previous studies of T cell reactivity to PMP-22 in either GBS or CIDP.
Evidence for T cell involvement in GBS and CIDP
Because of the improvement of patients after therapy with plasma exchange and intravenous immunoglobulin,20 antibody responses are thought to be important in GBS and CIDP. However, T cells are also likely to play a role in GBS and CIDP. T cell help is an important prerequisite for B cell maturation and antibody production. T cells are found in the peripheral nerves of some subjects with GBS21,22 and CIDP.23 Increased numbers of circulating activated T cells in patients with GBS,24,25 altered proportions of T cell subsets,25 raised circulating levels of IL-2 and soluble IL-2 receptor,26–28 and oligoclonal expansion of T cells bearing particular T cell receptor Vβ and Vδ genes29 all support a role for T cells in GBS and CIDP. Furthermore, experimental autoimmune neuritis (EAN), an animal model of GBS, which can be induced by inoculation of susceptible animals with myelin proteins, or peptides from P2,30 P0,31–34 or PMP-22,35 can also be passively transferred by activated T cells specific for peripheral myelin antigens.36
T cell Th1 versus Th2 cytokine secretion profiles
The spectrum of cytokines produced by T cells has a strong influence on the outcome of immune responses. Human CD4+ helper T cells can be divided into two categories. (a) Th1 cells produce the cytokines IL-2, IFN-γ, tumour necrosis factor (TNF)-α, and TNF-β. Th1 responses promote cell mediated inflammatory immune responses and are thought to be important for development of many organ specific autoimmune diseases.37,38 (b) Th2 type T cells produce IL-4, IL-5, IL-10, and IL-13, and are thought to suppress inflammation and help eosinophils and B cell antibody production, and may play beneficial roles in human organ specific inflammatory diseases.39 Th1 responses have been shown to be important in EAN induction, whereas the recovery phase is associated with a switch to Th2 responses.40,41
Overview of experiments
The importance of T cell reactivity to protein antigens of peripheral myelin in GBS and CIDP remains unclear. To define the frequency of these responses, we looked for proliferative T cell reactivity to peptides of P0, P2, MBP, and PMP-22 in a large group of patients with GBS and with CIDP, and compared these with responses of healthy controls and patients with ON.
We also used IFN-γ and IL-5 ELISPOT assays in a smaller subset of patients with GBS, CIDP, or ON and in healthy controls to characterise frequencies of both spontaneous and antigen specific cytokine secreting PBMC.
METHODS
Patients
The details of each patient group are shown in table 1. Patients with GBS, CIDP, or ON were recruited from local hospitals. Patients with GBS and those with CIDP fulfilled standard diagnostic criteria.42,43 Patients with GBS had predominantly demyelinating neuropathy and generally made a prompt recovery. Patients with Miller Fisher syndrome were excluded. Samples were collected from patients with GBS within 10 days of the onset of neurological symptoms, either before or at the peak of disease, and before administration of any treatment. Patients with GBS were graded on the GBS disability scale44 (table 2) and information regarding potential precipitating factors including recent infections or vaccinations was also recorded (table 2).
Patient number and age details, and proliferation assay background counts for patients with GBS, CIDP, or ON and healthy control subjects
GBS patient disease severity and potential precipitating factors
Patients with CIDP had not received any immunosuppressive or immunomodulatory treatment for 2 months prior to blood collection. For most of our patients with CIDP, blood was collected when they were hospitalised for treatment of relapses. Patients with ON included patients with hereditary motor sensory, toxic, and diabetic neuropathy. All blood samples were collected after informed, written consent was obtained from each subject.
Myelin proteins and peptides
MBP was purified from human brain tissue.45 Peptides used in the study were synthesised to >95% purity by Auspep (Melbourne, Australia). Table 3 summarises the characteristics of the antigens used in this study.
Peptide and protein antigens tested in this study, including sequences and numbering of residues within respective proteins
Proliferation assays
Proliferation assays using PBMC from each subject were prepared using methods previously outlined.47 Briefly, 105 PBMC were cultured as quadruplicates with and without test antigens (at 20 μg/ml) for 6 days, with 0.5 μCi 3H-thymidine being added during the last 18 hours of culture. Thymidine uptake was measured in counts per minute (cpm). The stimulation index (SI) was determined by the formula:
SI = (mean cpm of antigen containing wells)/(mean cpm of control wells)
A positive proliferative response was scored if the subject’s cells responded to the antigen tested with an SI ⩾2.0. Mean and standard error values for background counts for each subject group can be found in table 1.
ELISPOT assays
The numbers of spontaneous and antigen-specific IFN-γ and IL-5 secreting PBMC were determined using ELISPOT assays in 16 patients with GBS, 7 with CIDP, 13 with ON, and 14 healthy controls.
PBMC to be tested in ELISPOT assays were pre-incubated at 250 000 cells/well in 200 μl volumes of X-Vivo 15 media (Cambrex, NJ, USA) in the presence of test peptides or proteins at 20 μg/ml as quadruplicates, or without antigens as replicates of 12 wells, in 96 well, round bottomed, tissue culture plates for 18 hours to maximise cell–cell contact for antigen presentation. PBMC were then transferred to ELISPOT assay plates for the detection of human IFN-γ and IL-5 secretion. ELISPOT assays were conducted according to the manufacturer’s instructions (Mabtech, Sweden). The average number per well of spot forming cells (SFC) for each subject’s PBMC in the absence of antigen was calculated for both IFN-γ and IL-5, and for each subject this was used as a measure of spontaneous cytokine production and expressed as spontaneous SFC/106 PBMC. For each subject, and for each of the cytokines, the average spontaneous SFC was subtracted from the average SFC for PBMC in the presence of each of the test antigens, to produce a corrected SFC for each test antigen. Results were expressed as antigen specific SFC/106 PBMC.
We used IL-5 rather than IL-4 ELISPOT assays as a measure of T cell Th-2 cytokine secretion because IL-4 can be produced by cells other than T cells.48
Statistical analysis
Proliferation assays
In the different subject groups, the percentages of individuals making a positive proliferative response (SI ⩾2.0) to test peptides were compared using the χ2 test with Yates’ correction applied as required. Proliferative T cell reactivity to different antigens in the GBS or CIDP patient groups was deemed to be of significance only if p values for comparisons both with healthy controls and with patients with ON were ⩽0.05.
ELISPOT analysis of spontaneous and antigen specific IFN-γ and IL-5 secretion
ELISPOT data for both spontaneous and antigen specific IFN-γ and IL-5 secretion were found to be non-normally distributed. The means +2SD of spontaneous cytokine secretion and the means +2SD of antigen specific IFN-γ and IL-5 secretion in response to each antigen tested in the healthy control group were calculated and used as cutoffs for classifying data into positive and negative responses. This enabled spontaneous cytokine secretion levels and antigen specific reactivity within the four subject groups to each test antigen, and to all test antigens collectively grouped, to be compared by either χ2 test with Yates’ correction or Fishers exact test, and p⩽0.05 was deemed significant. Odds ratios (OR) with 95% confidence intervals (CI) were also calculated for significant comparisons.
RESULTS
T cell proliferation
The percentages of individuals in each subject group with a T cell reactivity SI ⩾2.0 were determined for each antigen (fig 1, table 4) and compared by χ2 analysis.
The percentages of individuals in each subject group with a T cell proliferation reactivity SI ⩾ 2.0 were determined for each peptide or protein and compared by χ2 analysis. Odds ratios (OR)with 95% confidence intervals (CI) were also calculated for selected comparisons

Percentages of patients with GBS, CIDP, or ON, and healthy controls (HC) responding with a proliferation assay SI ⩾2.0 to specific peptides and whole MBP.
The percentage of patients with GBS reacting with an SI ⩾2.0 to whole MBP (35.9%) was comparable with that seen in healthy controls (36.1%). The corresponding percentage of patients with CIDP (16.7%) was significantly lower than that of healthy controls (p = 0.013), but not significantly different from that of patients with ON. There was no statistically significant difference in reactivity to MBP82–100 between any of the test groups.
For PMP-2232–51, PMP-2241–61, and PMP-2251–64, no statistically significant differences in reactivity were seen between patients with GBS or CIDP, and healthy controls or patients with ON. The proportion of patients with GBS reacting to PMP-22120–133 (6.3%) was significantly lower than healthy controls (20.7%, p = 0.025), as was the proportion in the ON disease group (6.5%) to PMP-22120–133 (p = 0.031). Reactivity in patients with CIDP (13.2%) was also lower than that of healthy controls, although not significantly.
For P056–71 and P070–85, no statistically significant differences were seen between patients with GBS or CIDP, and healthy controls or patients with ON. The proportion of patients with GBS reacting to P0180–199 (19.6%) was significantly higher than that of healthy controls (6.5%, p = 0.042), but not higher than patients with ON (14.8%). Similarly, reactivity in patients with CIDP to P0180–199 (18.8%) was higher than that of healthy controls, but not significantly (p = 0.068).
No significant differences in proliferative reactivity to either P214–25 or P261–70 were seen between patients with GBS or CIDP, and healthy controls or patients with ON. When GBS patient reactivity to either P214–25 or P261–70 (19%) was compared with the reactivity of healthy controls and patients with ON grouped collectively (9%), significance was reached (p = 0.029). Similarly, reactivity of patients with CIDP to either of these peptides (20.7%) was significantly greater than that of healthy controls and patients with ON grouped collectively (p = 0.018).
ELISPOT analysis
ELISPOT analysis of spontaneous IFN-γ and IL-5 secretion
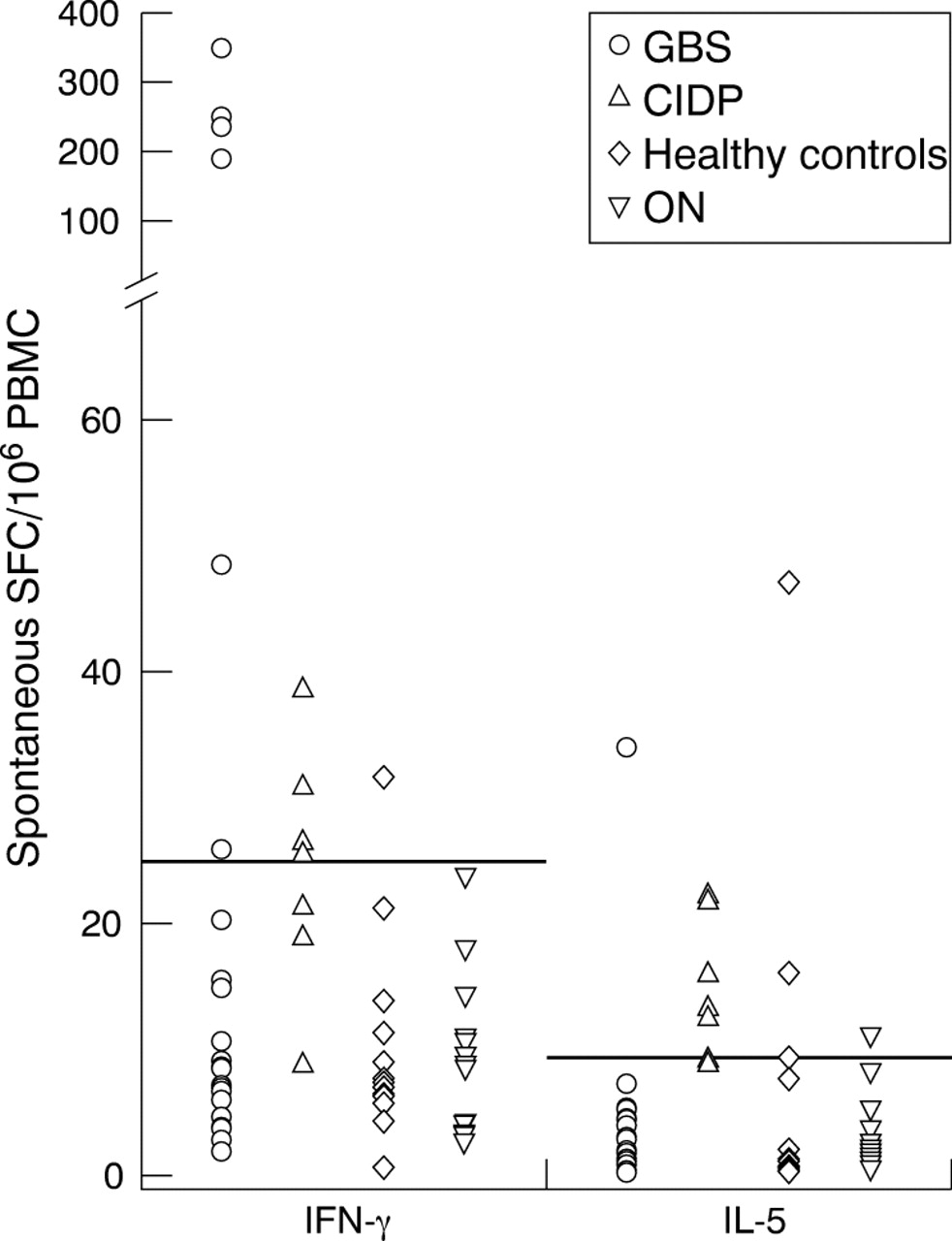
Numbers of ex vivo, spontaneous (in the absence of antigen) IFN-γ and IL-5 secreting PBMC were enumerated using ELISPOT assays in 24 patients with GBS, 7 with CIDP, 13 with ON, and 14 healthy controls (fig 2).

Scatter plot of the numbers of spontaneous IFN-γ and IL-5 secreting PBMC in patients with GBS, CIDP, or ON, and in healthy controls. The short horizontal lines represent the threshold cutoffs for positive responses. Results are expressed as spot forming cells (SFC) per 106 PBMC.
The proportion of patients with CIDP with an increased number of PBMC spontaneously secreting IFN-γ (57.1%) was significantly higher than that in healthy controls alone (7.1%, p = 0.025), patients with ON alone (0%, p = 0.002), and in grouped healthy controls and patients with ON (3.7%, p = 0.005, odds ratio (OR) = 34.6, 95% confidence interval (CI) 2.8 to 420.2). The proportion of patients with GBS with an increased number of PBMC spontaneously secreting IFN-γ (25.0%) was significantly higher than in grouped healthy controls plus patients with ON (p = 0.042, OR = 8.7, 95% CI 1.9 to 78.3), and in patients with ON alone (p = 0.048), but bordered upon significance when compared with healthy controls alone (p = 0.07). Interestingly, the highest values of spontaneous IFN-γ secretion of all responses tested were seen in five of the 24 patients with GBS tested.
The proportion of subjects with an increased number of PBMC spontaneously secreting IL-5 was significantly higher in patients with CIDP, but not those with GBS, compared with healthy controls (p = 0.048) and patients with ON (p = 0.008).
ELISPOT analysis of antigen specific IFN-γ and IL-5 secretion
Numbers of ex vivo, background corrected, antigen specific IFN-γ and IL-5 secreting PBMC were determined using ELISPOT assays in 16 patients with GBS, 7 with CIDP, 13 with ON, and 14 healthy controls (fig 3A, B). The proportions of subjects in each of the four groups, and also in the combined group of healthy controls and patients with ON, with an increased number of PBMC producing IFN-γ and IL-5 in response to stimulation with individual peptides and whole MBP are shown in table 5.
Percentages of patients with GBS, CIDP, or ON and healthy controls (HC) responding with positive antigen specific ELISPOT IFN-γ and IL-5 reactivity to specific peptides and whole MBP

{kind=link}
{kind=link}
{kind=link}
Scatter plot of antigen specific (A) IFN-γ and (B) IL-5 secretion after stimulation of PBMC with each of the test antigens in patients with GBS, CIDP, or ON, and healthy controls (HC). Background numbers of PBMC spontaneously producing IFN-γ or IL-5 in the absence of antigen have been subtracted for each data point. The short horizontal lines represent the threshold cutoffs for positive responses to each antigen tested. Results are expressed as spot forming cells (SFC) per 106 PBMC. For sequences of peptides tested, refer to table 3.
Antigen specific ELISPOT data for each cytokine were initially analysed for each individual test antigen. Reactivity to each antigen in patients with either GBS or CIDP was compared with healthy controls alone, with patients with ON alone, and with collectively grouped healthy controls and patients with ON. The proportion of patients with GBS with an increased number of PBMC producing IFN-γ in response to peptide P070–85 (25.0%) was significantly higher than grouped healthy controls plus patients with ON (0%, p = 0.027, OR = 9.0, 95% CI 0.90 to 82.6), but only approached significance when compared with healthy controls alone (p = 0.066) and patients with ON alone (p = 0.077). The proportion of patients with CIDP with increased numbers of PBMC producing IFN-γ in response to peptide PMP-2251–64 (57.1%) was significantly higher than both healthy controls alone (p = 0.025, OR = 17.33, 95% CI 1.38 to 216.6) and patients with ON alone (p = 0.007, OR = 18.66, 95% CI 1.50 to 232.3).
The proportion of patients with CIDP with increased numbers of PBMC producing IFN-γ in response to peptide PMP-22120–133 (42.8 %) was significantly higher than that of grouped healthy controls plus patients with ON (3.7%) (p = 0.021, OR = 19.5, 95% CI 1.60 to 236.6), and of patients with ON alone (p = 0.031), but not that of healthy controls alone (p = 0.088).
No other significant differences in antigen specific IFN-γ production to any of the other individual peptides or whole MBP were noted in either patients with GBS or patients with CIDP compared with the grouped healthy controls and patients with ON. No significant differences in antigen specific IL-5 secretion were evident between patients with either GBS or CIDP and the grouped healthy controls and patients with ON using χ2 analysis for comparison of reactivity to individual test peptides.
As a second method for statistically analysing antigen specific ELISPOT results, the GBS patient reactivity data for all antigens tested were grouped into a combined dataset, as were data for patients with CIDP or ON and healthy controls. Analysis of this grouped data showed a significantly increased proportion of antigen specific IFN-γ secretion in patients with GBS compared with healthy controls (p = 0.03, OR = 2.30, 95% CI 1.02 to 5.16) and patients with ON (p = 0.03, OR = 2.41, 95% CI 1.03 to 5.59). Similarly, antigen specific IFN-γ secretion in patients with CIDP was significantly increased compared with healthy controls (p = 0.0000006, OR = 6.8, 95% CI 2.98 to 15.76) and with patients with ON (p = 0.00000079, OR = 7.18, 95% CI 3.02 to 17.05). No significant differences in antigen specific IL-5 secretion between either patients with GBS or patients with CIDP and healthy controls or patients with ON were evident using this grouped data analysis (fig 3B, table 5).
It is important to note that in any study involving a large number of formal statistical tests, on average 1 in 20 comparisons will reach a significant p value of 0.05 by chance.
Correlation between peptide reactivity and GBS disease severity
Patients with GBS were grouped into positive and negative responders for antigen specific proliferation to peptides derived from each peripheral myelin protein, and for reactivity to any of the peptides tested. Mean GBS disability scale values were calculated within each of the subgroups (table 6). Interestingly, the subgroup of patients with GBS with positive T cell reactivity to any of the PMP-22 peptides had a statistically lower GBS disability score than the subgroup of non-responding patients (p = 0.003, t test). Disability assessments were not made for patients with CIDP at the time of blood collection, so no correlations could be made for CIDP.
Correlations between GBS patient T cell peptide reactivity and disease severity
In patients with GBS, no statistically significant correlation could be made linking reactivity to any of the antigens tested with potential precipitating factors such as type of antecedent infection.
DISCUSSION
In patients with GBS, CIDP, or ON and healthy controls, we performed a detailed comparative study of T cell proliferative reactivity and antigen specific ELISPOT IFN-γ and IL-5 secretion in response to antigenic stimulation with peptides derived from P0, P2, PMP-22, and MBP. We also compared the numbers of PBMC spontaneously secreting IFN-γ and IL-5 in patients with GBS or CIDP with those in healthy controls and patients with ON.
Proliferation assay reactivity
No statistically significant elevations in frequencies of antigen specific proliferation assay reactivity to any of the antigens tested were seen in patients with GBS or CIDP compared with both healthy controls and patients with ON. The proportion of patients with GBS reacting to P0180–199 was significantly higher than that of healthy controls, but not patients with ON. There was some increase in reactivity to P2 and P0 peptides in both patients with GBS and those with CIDP, but this was not statistically significant.
This study was not an exhaustive search for T cell epitope specificity within peripheral myelin proteins. There may be T cells of pathological importance in GBS and CIDP specific to other epitopes within P0, P2, and PMP-22 that were not encompassed by the peptides selected for study in this project, or specific for other peripheral nerve myelin antigens. We previously found significantly increased PBMC proliferative reactivity to the ganglioside GM1 in patients with GBS,49 and it may be that glycolipid antigens also serve as targets of autoreactivity.
Interestingly, we found that patients with GBS with positive proliferative reactivity to PMP-22 peptides as a group had significantly less severe disease than patients not responding to these peptides. We previously found a similar correlation, in that patients with GBS with anti-GM1 T cell reactivity were significantly less likely to need mechanical ventilation than those without such reactivity.49 This raises the possibility that T cells specific for certain peripheral nerve components, such as PMP-22 and GM1, might have an immunoregulatory or neuroprotective role.
Th1 versus Th2 cytokines in GBS
In patients with GBS, we found significantly increased numbers of PBMC spontaneously secreting IFN-γ compared with grouped healthy controls plus patients with ON, and to patients with ON alone. Although comparisons with healthy controls alone only bordered upon significance, the highest values of spontaneous IFN-γ secretion were seen in five of the 24 tested patients with GBS. These cells are presumably reacting to in vivo stimulation with unknown peptide or ganglioside antigens. They may be contributing to the immunopathogenesis of GBS in some patients, or may persist as a consequence of T cell activation by the antecedent infections which commonly precede GBS.4,5 It may be that T cells secreting pro-inflammatory Th1 type cytokines are important in the early stages of disease in some patients with GBS, possibly reducing in numbers by the time neurological symptoms are evident. Blood was collected from patients in our study soon after the onset of neurological symptoms, sometimes within 2 days.
In patients with GBS, we found a significantly higher proportion of subjects with an increased number of PBMC secreting IFN-γ in response to all test antigens grouped collectively compared with healthy controls alone and patients with ON alone, and in response to P070–85 compared with grouped healthy controls plus patients with ON, but none of the responses to individual test antigens in patients with GBS was significantly higher than in both healthy controls alone and in patients with ON alone.
There are conflicting reports of elevated levels of spontaneous and antigen specific Th1 and Th2 cytokine secreting cells in early GBS,50–53 and several studies outline an increasing involvement for Th2 cytokine secreting cells in the recovery stages of GBS.54,55 It has been postulated that IL-4 (Th2) responses are beneficial in GBS, and may have a role in disease termination through downregulation of Th1 responses.39,54 Conversely, Th2 cytokines may induce allergic disease by helping B cells produce potentially pathogenic allergen specific IgE,51 which is reportedly increased in both GBS and CIDP.56 GBS is usually a self limiting disease, and Th2 cytokine production has been reported in other diseases that resolve spontaneously such as reactive arthritis.57 However, in our study, both antigen specific and spontaneous levels of IL-5 secretion in patients with GBS did not differ significantly from healthy controls and patients with ON. We tested GBS subjects only in the early stage of disease, so we were unable to enumerate Th2 cytokine producing cells in the recovery phase of GBS.
Th1 versus Th2 cytokines in CIDP
We have shown significantly increased numbers of PBMC spontaneously secreting IFN-γ and IL-5 in patients with CIDP. We also found a significantly higher proportion of patients with CIDP with increased numbers of PBMC secreting IFN-γ in response to PMP-2251–64 and in response to all test antigens grouped collectively compared with healthy controls alone and patients with ON alone. Of the four patients with CIDP with increased numbers of PBMC secreting IFN-γ in response to PMP-2251–64, only one showed positive proliferative reactivity to this peptide, emphasising the increased detection sensitivity of ELISPOT IFN-γ assays.
For many of our patients with CIDP, blood was collected when they were hospitalised for treatment of active disease exacerbation. A difference in cytokine profiles between the exacerbation and remission stages of CIDP has been reported.58 Our finding of increased numbers of spontaneous Th1 IFN-γ and Th2 IL-5 secreting cells in patients with CIDP is partly at odds with a previous report of increased proportions of only Th2-IL-4 secreting CD4+T cells in the peripheral blood of 12 patients with CIDP.51 We found increased numbers of spontaneous, but not antigen specific, IL-5 secreting PBMC in patients with CIDP compared with healthy controls and patients with ON. Others have found increased levels of both the Th1 (IFN-γ) and Th2 (IL-4) cytokines in the exacerbation stage of CIDP, but an elevation of only IL-4 secreting Th2 cells in remission.58 It is important to note that our ELISPOT methodology detected IL-5 rather than IL-4 as a measure of Th2 cytokine secretion. CIDP patient cells tested in ELISPOT assays were all collected during exacerbations, so we were unable to assess numbers of Th1 and Th2 cytokine producing cells during remission.
Measurements of both spontaneous and antigen specific cytokine secreting cells in the peripheral blood may under-represent the importance of these cells in both GBS and CIDP. Their numbers in the peripheral blood may be reduced because of homing to the peripheral nervous system, perhaps even before the onset of neurological symptoms.
We have shown that T cells specific for peripheral myelin peptides are present in the blood of patients with CIDP at low frequencies. It may be possible that these low levels of T cells, upon crossing the blood–nerve barrier and coming into contact with their respective antigens in the endoneural compartment, are further activated to proliferate and produce pro-inflammatory cytokines and chemokines. This may then be sufficient to increase blood–nerve barrier permeability to antibody,59 with resultant complement fixation and other pro-inflammatory immunological processes.
CONCLUSIONS
Our findings of increased numbers of spontaneous IFN-γ secreting PBMC in 25% of patients with GBS suggests that Th1 cells may play a role in the pro-inflammatory disease process in early GBS, although the specificity of these cells remains to be determined. Significantly increased levels of both spontaneous and antigen specific IFN-γ secretion, as well as increased spontaneous IL-5 secretion in patients with CIDP, suggest that T cells present in the blood of patients with CIDP have both pro-inflammatory and immunoregulatory potential.
Although significantly increased numbers of PMP-2251–64 specific IFN-γ secreting PBMC were found in patients with CIDP, the frequencies of these cells were generally lower than 1 in 10 000 cells, and this low level reactivity was not detected in proliferation assays.
Further careful analysis of T cell epitopic specificity and Th1/Th2 cytokine secretion profiles in the blood from early disease through to recovery in patients with GBS, and in exacerbations and remissions of CIDP are required to shed further light on the roles of peripheral myelin reactive T cells in these diseases.
Acknowledgments
We wish to acknowledge the Wesley Research Institute and the National Health and Medical Research Council of Australia for support and research funding, the Australian Red Cross Blood Transfusion Service for supplying human serum as a tissue culture additive, and the Commonwealth Serum Laboratories (Melbourne, Australia) for providing tetanus toxoid.
REFERENCES
Footnotes
-
Competing interests: none declared
-
Ethics approval: This study was approved by the Human Research Ethics committees of the Royal Brisbane, Princess Alexandra, Mater, Greenslopes Private, and Logan Hospitals, as well as the Medical Research Ethics committee of the University of Queensland.